

Inflammatory factors of the tumor microenvironment induce plasticity in nontransformed breast epithelial cells: EMT, invasion, and collapse of normally organized breast textures
- PMID: 24403855
- PMCID: PMC3884524
- DOI: 10.1593/neo.131688
Inflammatory factors of the tumor microenvironment induce plasticity in nontransformed breast epithelial cells: EMT, invasion, and collapse of normally organized breast textures
Abstract
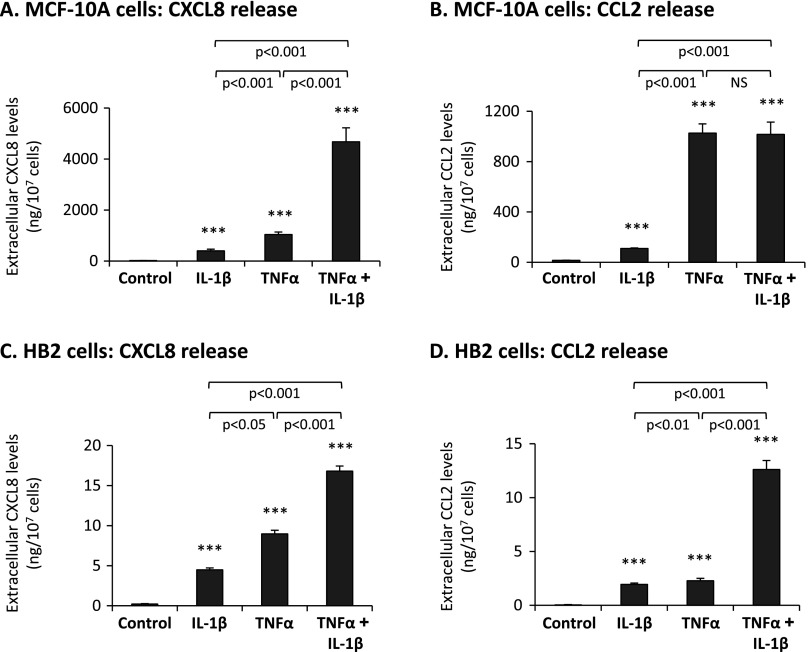
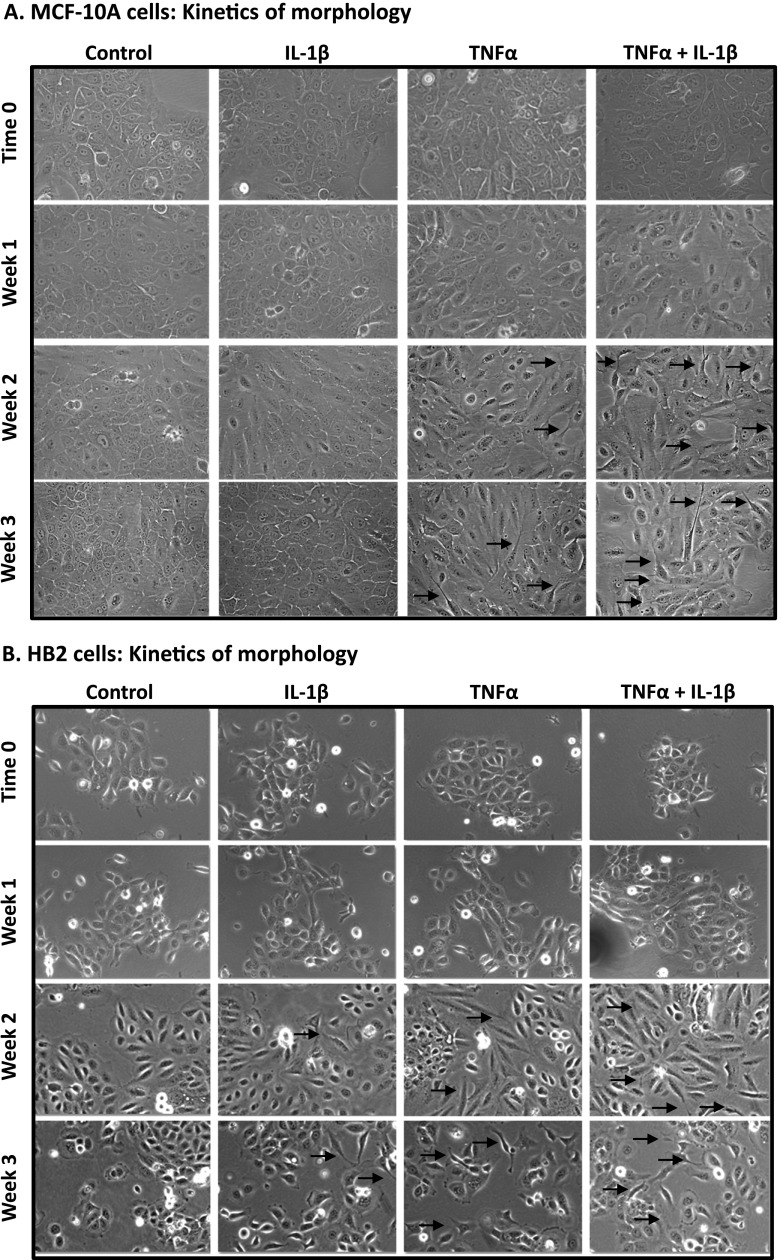
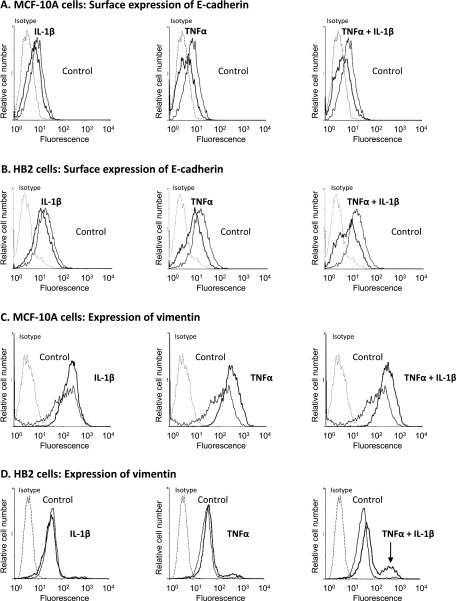
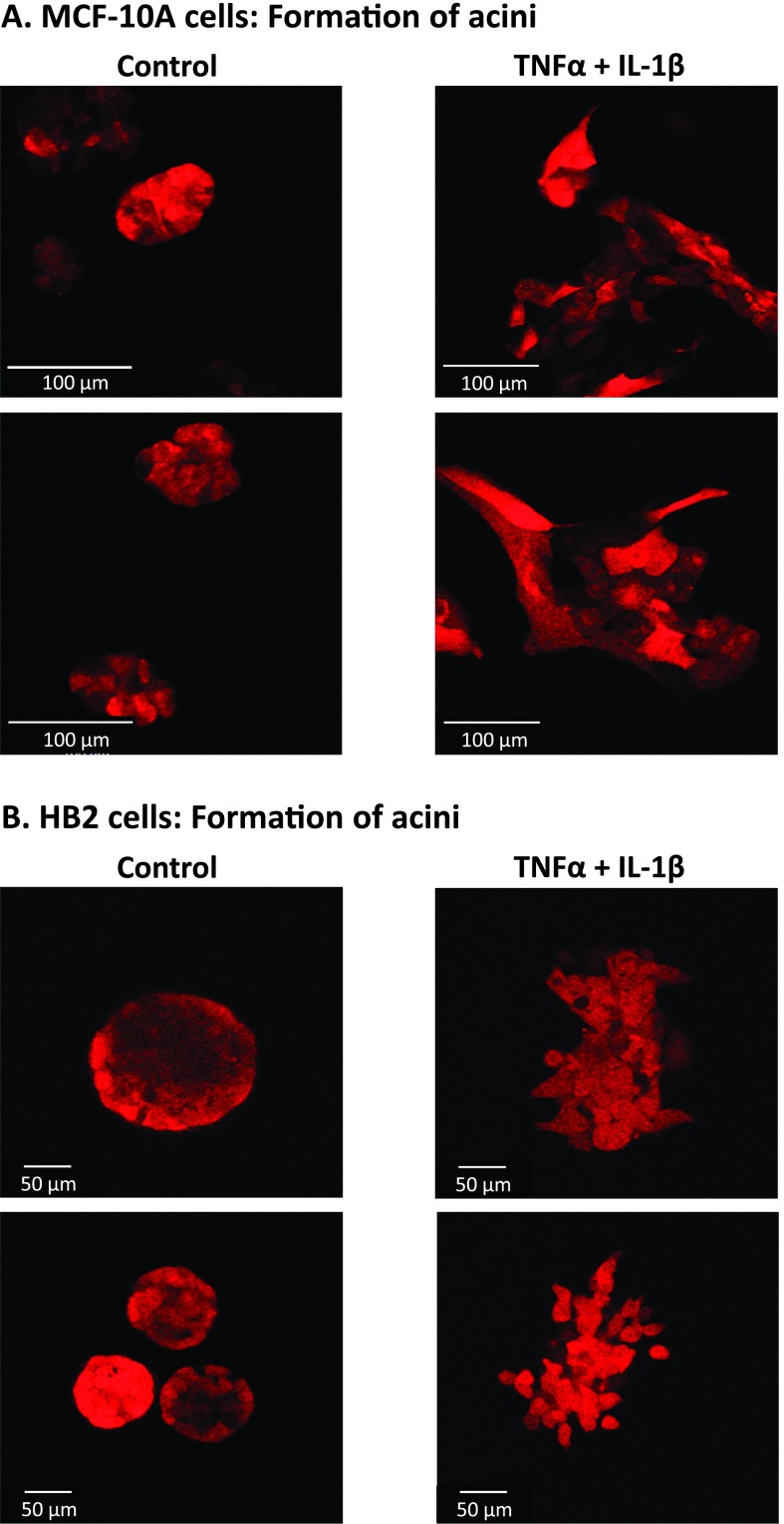
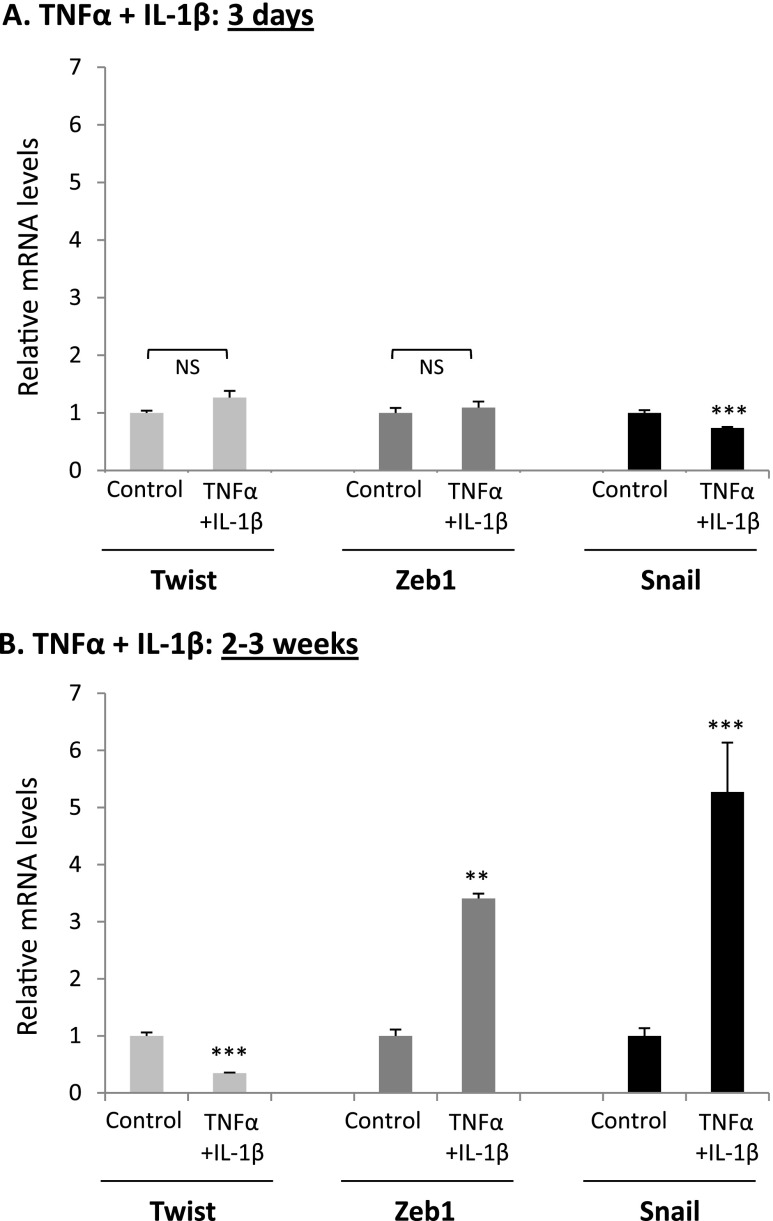
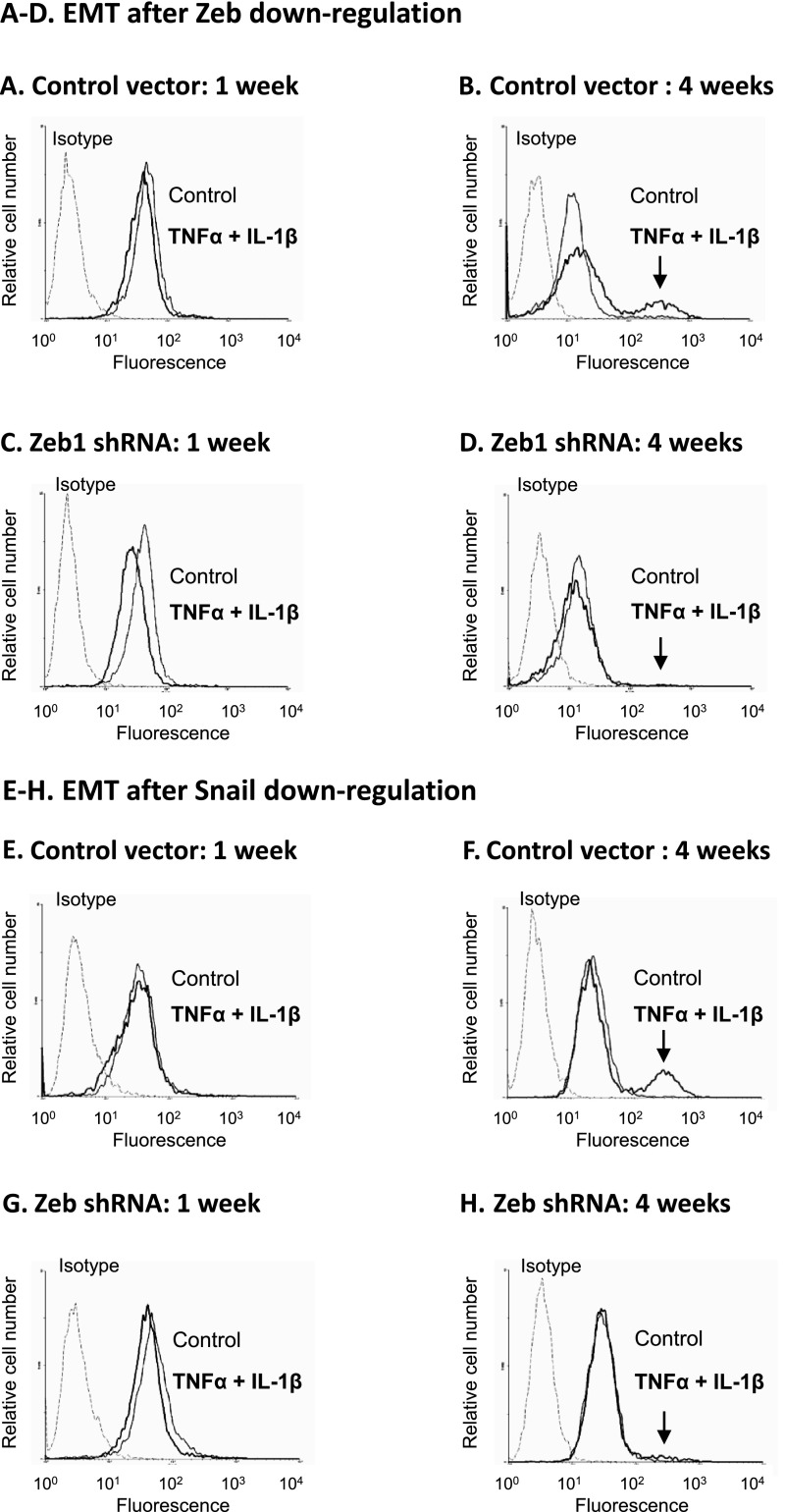
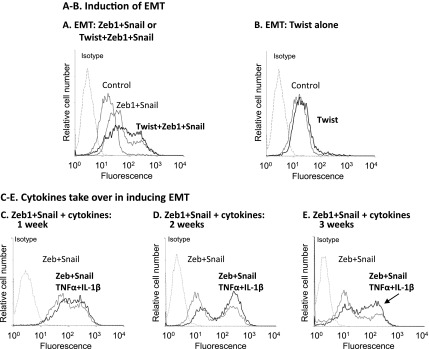
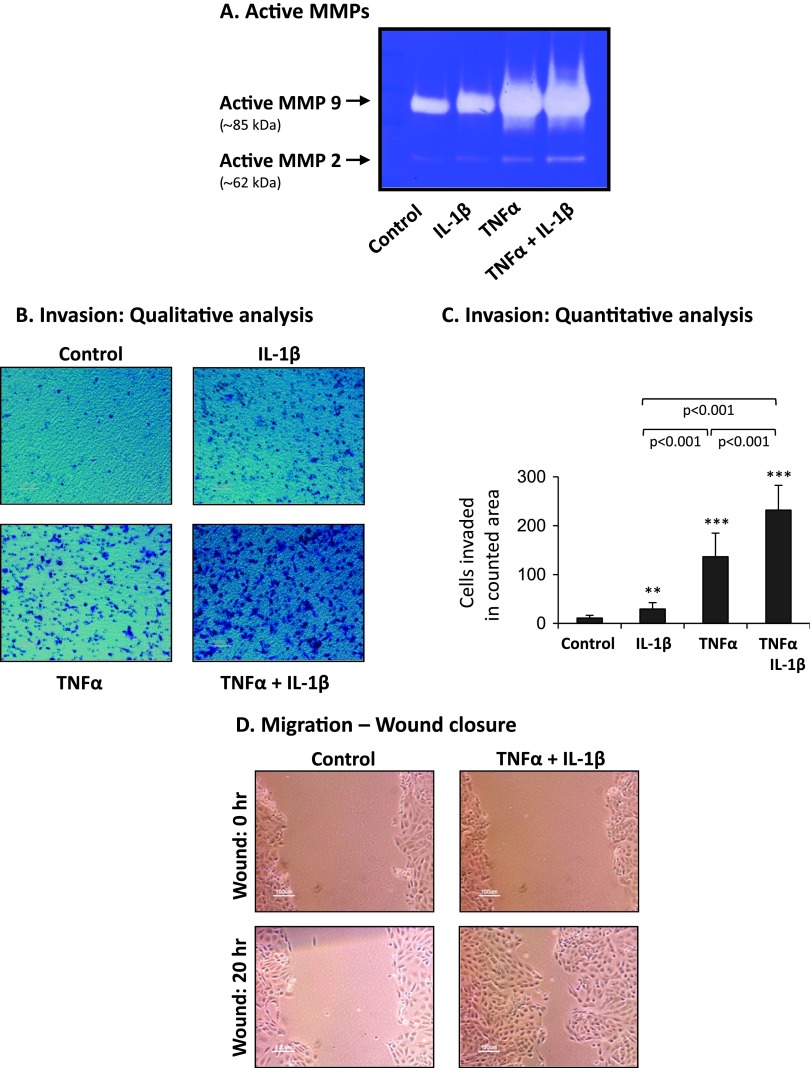
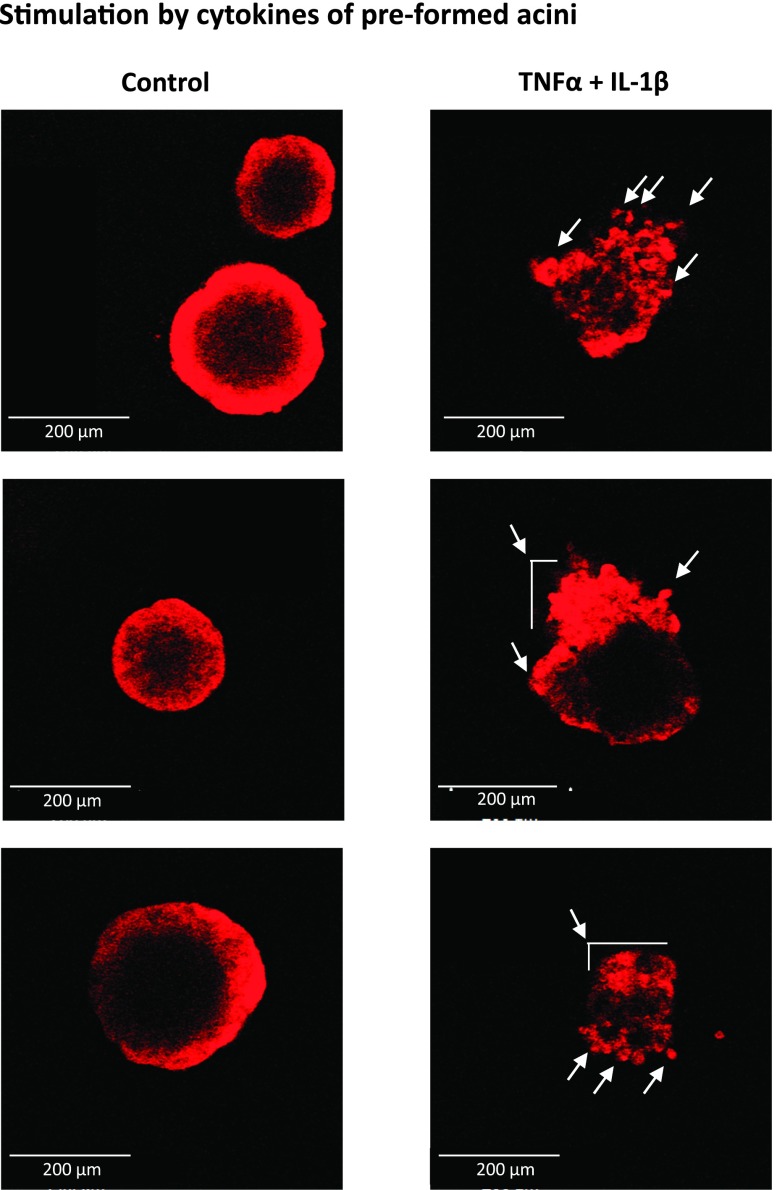
Nontransformed breast epithelial cells that are adjacent to tumor cells are constantly exposed to tumor necrosis factor-α (TNFα) and interleukin-1β (IL-1β), two inflammatory cytokines identified as having pro-tumoral causative roles. We show that continuous stimulation of nontransformed breast epithelial cells by TNFα + IL-1β for 2 to 3 weeks induced their spreading and epithelial-to-mesenchymal transition (EMT). The mechanistic bases for this slow induction of EMT by TNFα + IL-1β are: 1) it took 2 to 3 weeks for the cytokines to induce the expression of the EMT activators Zeb1 and Snail; 2) although Twist has amplified the EMT-inducing activities of Zeb1 + Snail, its expression was reduced by TNFα + IL-1β; however, the lack of Twist was compensated by prolonged stimulation with TNFα + IL-1β that has potentiated the EMT-inducing activities of Zeb1 + Snail. Stimulation by TNFα + IL-1β has induced the following dissemination-related properties in the nontransformed cells: 1) up-regulation of functional matrix metalloproteinases; 2) induction of migratory and invasive capabilities; 3) disruption of the normal phenotype of organized three-dimensional acini structures typically formed only by nontransformed breast cells and spreading of nontransformed cells out of such acini. Our findings suggest that TNFα + IL-1β induce dissemination of nontransformed breast epithelial cells and their reseeding at the primary tumor site; if, then, such detached cells are exposed to transforming events, they may form secondary malignant focus and lead to disease recurrence. Thus, our study reveals novel pathways through which the inflammatory microenvironment may contribute to relapsed disease in breast cancer.
Figures
Similar articles
-
Evidence that GTP-binding domain but not catalytic domain of transglutaminase 2 is essential for epithelial-to-mesenchymal transition in mammary epithelial cells.Breast Cancer Res. 2012 Jan 6;14(1):R4. doi: 10.1186/bcr3085. Breast Cancer Res. 2012. PMID: 22225906 Free PMC article.
-
Inflammatory mediators in breast cancer: coordinated expression of TNFα & IL-1β with CCL2 & CCL5 and effects on epithelial-to-mesenchymal transition.BMC Cancer. 2011 Apr 12;11:130. doi: 10.1186/1471-2407-11-130. BMC Cancer. 2011. PMID: 21486440 Free PMC article.
-
IL-1β promotes stemness and invasiveness of colon cancer cells through Zeb1 activation.Mol Cancer. 2012 Nov 23;11:87. doi: 10.1186/1476-4598-11-87. Mol Cancer. 2012. PMID: 23174018 Free PMC article.
-
Exploring the Crosstalk between Inflammation and Epithelial-Mesenchymal Transition in Cancer.Mediators Inflamm. 2021 Jun 14;2021:9918379. doi: 10.1155/2021/9918379. eCollection 2021. Mediators Inflamm. 2021. PMID: 34220337 Free PMC article. Review.
-
Oxidative stress regulation and related metabolic pathways in epithelial-mesenchymal transition of breast cancer stem cells.Stem Cell Res Ther. 2023 Nov 28;14(1):342. doi: 10.1186/s13287-023-03571-6. Stem Cell Res Ther. 2023. PMID: 38017510 Free PMC article. Review.
Cited by
-
[Research progress on epithelial-mesenchymal transition in cancer recurrence and metastasis].Zhejiang Da Xue Xue Bao Yi Xue Ban. 2015 Mar;44(2):211-6. doi: 10.3785/j.issn.1008-9292.2015.03.015. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2015. PMID: 26038142 Free PMC article. Review. Chinese.
-
Chemokine-Derived Peptides: Novel Antimicrobial and Antineoplasic Agents.Int J Mol Sci. 2015 Jun 8;16(6):12958-85. doi: 10.3390/ijms160612958. Int J Mol Sci. 2015. PMID: 26062132 Free PMC article. Review.
-
Activation of platelet protease-activated receptor-1 induces epithelial-mesenchymal transition and chemotaxis of colon cancer cell line SW620.Oncol Rep. 2015 Jun;33(6):2681-8. doi: 10.3892/or.2015.3897. Epub 2015 Apr 3. Oncol Rep. 2015. PMID: 25846512 Free PMC article.
-
Importance of EMT Factor ZEB1 in cDC1 "MutuDC Line" Mediated Induction of Th1 Immune Response.Front Immunol. 2018 Nov 13;9:2604. doi: 10.3389/fimmu.2018.02604. eCollection 2018. Front Immunol. 2018. PMID: 30483264 Free PMC article.
-
Mesenchymal traits at the convergence of tumor-intrinsic and -extrinsic mechanisms of resistance to immune checkpoint blockers.Emerg Top Life Sci. 2017 Dec 12;1(5):471-486. doi: 10.1042/ETLS20170068. Emerg Top Life Sci. 2017. PMID: 33525801 Free PMC article.
References
-
- Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A. Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis. 2009;30:1073–1081. - PubMed
-
- Allavena P, Sica A, Solinas G, Porta C, Mantovani A. The inflammatory micro-environment in tumor progression: the role of tumor-associated macrophages. Crit Rev Oncol Hematol. 2007;66:1–9. - PubMed
-
- Goldberg JE, Schwertfeger KL. Proinflammatory cytokines in breast cancer: mechanisms of action and potential targets for therapeutics. Curr Drug Targets. 2013;11:1133–1146. - PubMed
-
- Jin L, Yuan RQ, Fuchs A, Yao Y, Joseph A, Schwall R, Schnitt SJ, Guida A, Hastings HM, Andres J, et al. Expression of interleukin-1beta in human breast carcinoma. Cancer. 1997;80:421–434. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials