

Characterization of T cell phenotype and function in a double transgenic (collagen-specific TCR/HLA-DR1) humanized model of arthritis
- PMID: 24405551
- PMCID: PMC3978884
- DOI: 10.1186/ar4433
Characterization of T cell phenotype and function in a double transgenic (collagen-specific TCR/HLA-DR1) humanized model of arthritis
Abstract
Introduction: T cells orchestrate joint inflammation in rheumatoid arthritis (RA), yet they are difficult to study due to the small numbers of antigen-specific cells. The goal of this study was to characterize a new humanized model of autoimmune arthritis and to describe the phenotypic and functional changes that occur in autoimmune T cells following the induction of pathological events.
Methods: We developed a double transgenic mouse containing both the HLA-DR1 transgene and an HLA-DR1-restricted collagen-specific TCR in order to obtain large numbers of antigen-specific T cells that can be used for immunologic studies.
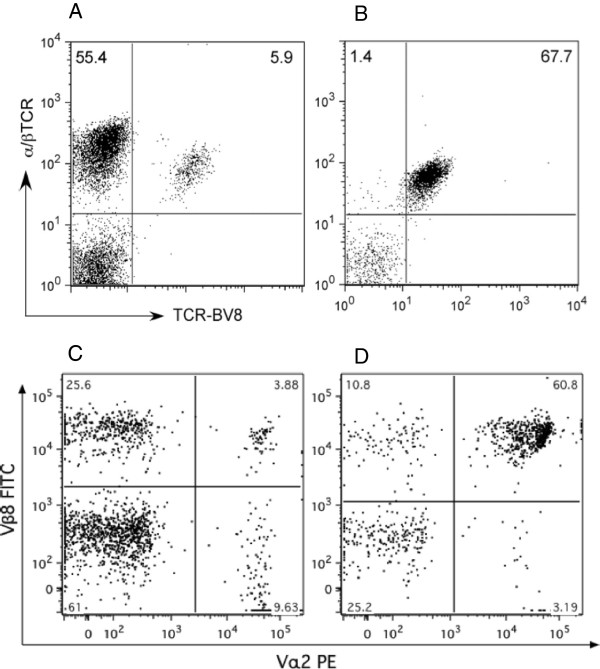
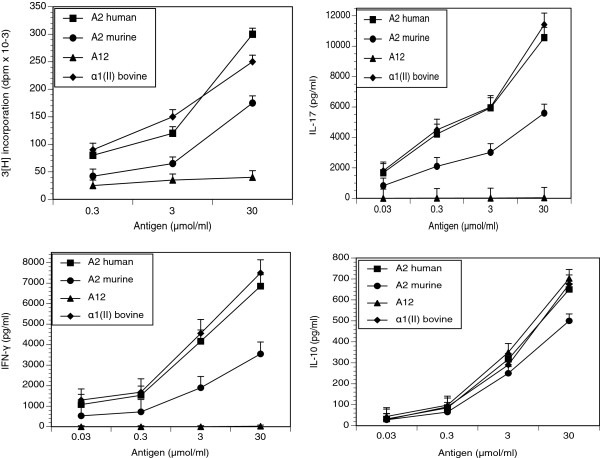
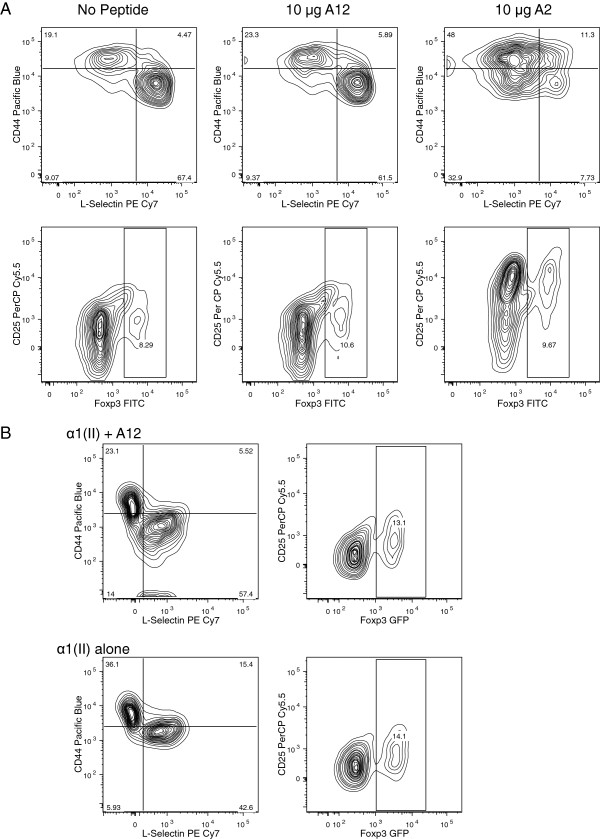
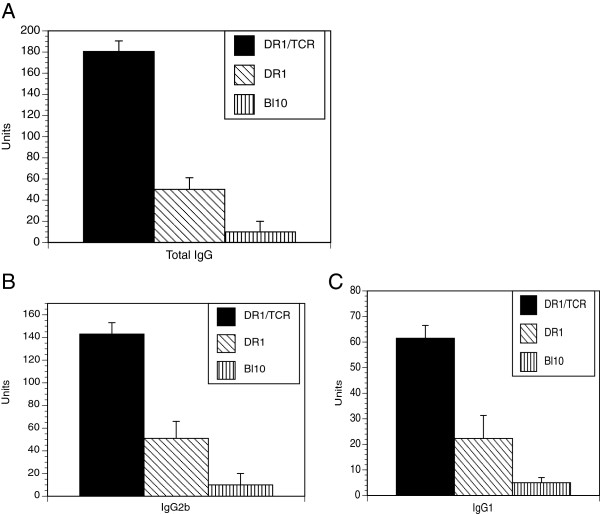
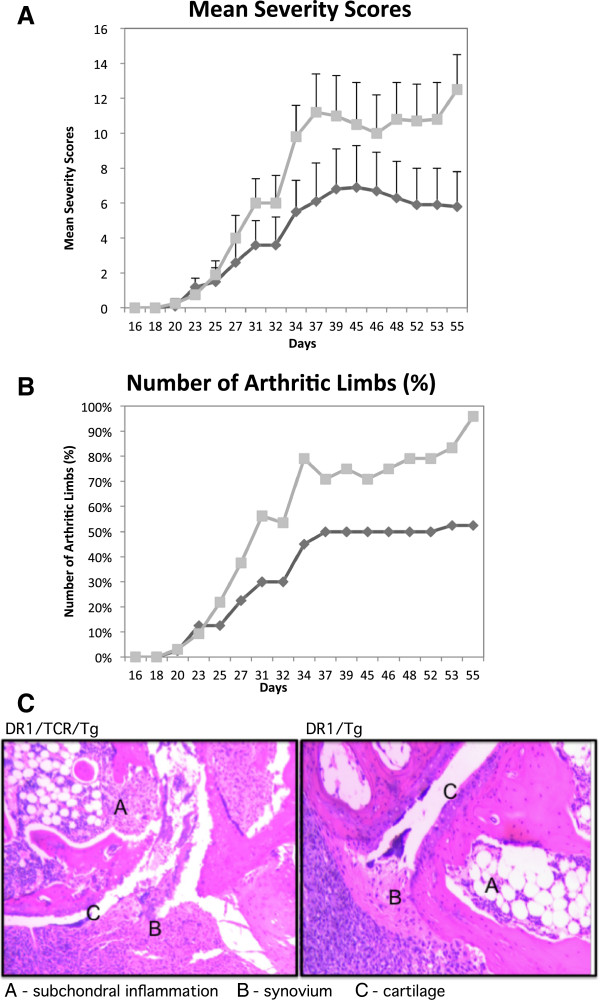
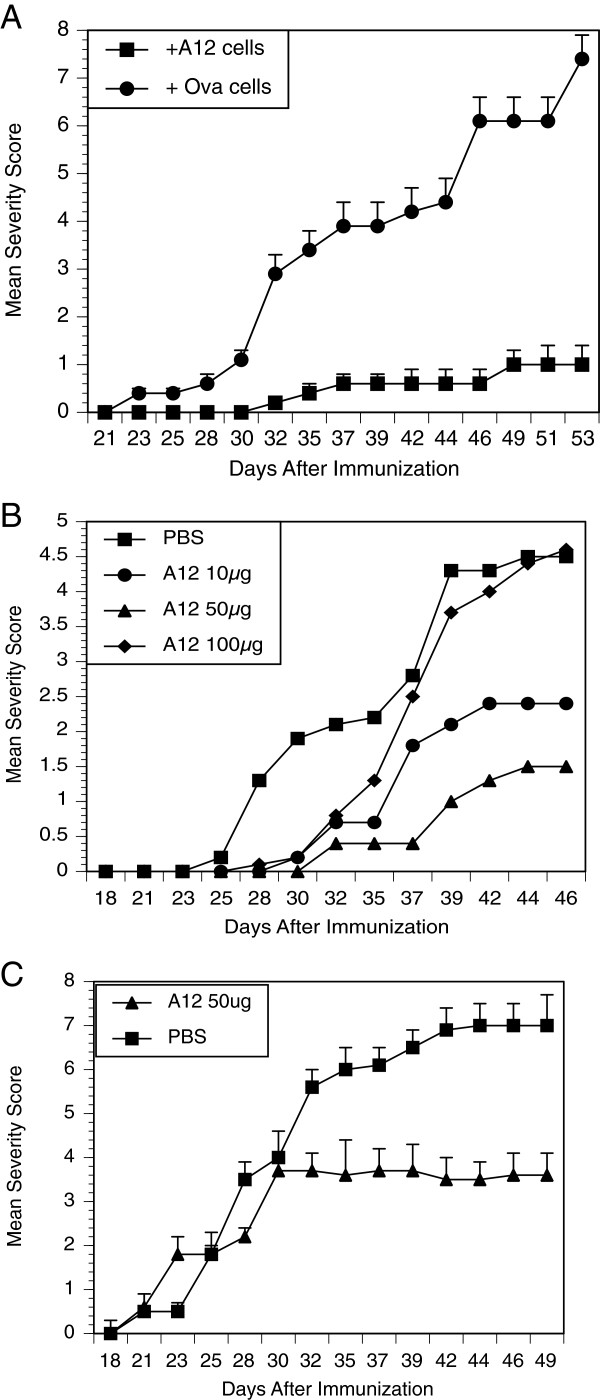
Results: In vitro, CII-specific T cells from this mouse proliferated vigorously in response to the CII immunodominant peptide A2 and the cells altered their phenotype to become predominately CD62Llow and CD44high "activated" T cells. The response was accompanied by the production of Th1, Th2, and Th17-type cytokines. Following immunization with bovine CII/CFA, these mice develop an accelerated arthritis compared to single transgenic HLA-DR1 mice. On the other hand, when the mice were treated orally with the analog peptide A12, (a suppressive analog of collagen we have previously described), arthritis was significantly suppressed, despite the fact that >90% of the CD4+ T cells express the TCR Tg. In GALT tissues taken from the A12-treated mice, IL-2, IFN-γ, and IL-17 production to the autoimmune collagen determinant dropped while high levels of IL-10 and IL-4 were produced.
Conclusions: We have developed a humanized model of autoimmune arthritis that will be useful for the study of T cell directed therapies as well as T cell mediated mechanisms of autoimmune diseases.
Figures
Similar articles
-
CD8+ T Cells Expressing an HLA-DR1 Chimeric Antigen Receptor Target Autoimmune CD4+ T Cells in an Antigen-Specific Manner and Inhibit the Development of Autoimmune Arthritis.J Immunol. 2022 Jan 1;208(1):16-26. doi: 10.4049/jimmunol.2100643. Epub 2021 Nov 24. J Immunol. 2022. PMID: 34819392 Free PMC article.
-
Characterization of inhibitory T cells induced by an analog of type II collagen in an HLA-DR1 humanized mouse model of autoimmune arthritis.Arthritis Res Ther. 2012 May 8;14(3):R107. doi: 10.1186/ar3832. Arthritis Res Ther. 2012. PMID: 22569209 Free PMC article.
-
Ex vivo characterization of the autoimmune T cell response in the HLA-DR1 mouse model of collagen-induced arthritis reveals long-term activation of type II collagen-specific cells and their presence in arthritic joints.J Immunol. 2005 Apr 1;174(7):3978-85. doi: 10.4049/jimmunol.174.7.3978. J Immunol. 2005. PMID: 15778354
-
Immunopathogenesis of collagen arthritis.Springer Semin Immunopathol. 2003 Aug;25(1):3-18. doi: 10.1007/s00281-003-0127-1. Springer Semin Immunopathol. 2003. PMID: 12904888 Review.
-
Transgenic mouse models of rheumatoid arthritis.Immunol Rev. 1999 Jun;169:161-73. doi: 10.1111/j.1600-065x.1999.tb01314.x. Immunol Rev. 1999. PMID: 10450516 Review.
Cited by
-
Role of the neutrophil chemorepellent soluble dipeptidyl peptidase IV in decreasing inflammation in a murine model of arthritis.Arthritis Rheumatol. 2015 Oct;67(10):2634-8. doi: 10.1002/art.39250. Arthritis Rheumatol. 2015. PMID: 26138693 Free PMC article.
-
Humanized Mouse Models of Rheumatoid Arthritis for Studies on Immunopathogenesis and Preclinical Testing of Cell-Based Therapies.Front Immunol. 2019 Feb 19;10:203. doi: 10.3389/fimmu.2019.00203. eCollection 2019. Front Immunol. 2019. PMID: 30837986 Free PMC article. Review.
-
CD8+ T Cells Expressing an HLA-DR1 Chimeric Antigen Receptor Target Autoimmune CD4+ T Cells in an Antigen-Specific Manner and Inhibit the Development of Autoimmune Arthritis.J Immunol. 2022 Jan 1;208(1):16-26. doi: 10.4049/jimmunol.2100643. Epub 2021 Nov 24. J Immunol. 2022. PMID: 34819392 Free PMC article.
-
1,25-Dihydroxyvitamin D3 and 20-Hydroxyvitamin D3 Upregulate LAIR-1 and Attenuate Collagen Induced Arthritis.Int J Mol Sci. 2021 Dec 12;22(24):13342. doi: 10.3390/ijms222413342. Int J Mol Sci. 2021. PMID: 34948139 Free PMC article.
-
Ameliorating effects of Gö6976, a pharmacological agent that inhibits protein kinase D, on collagen-induced arthritis.PLoS One. 2019 Dec 6;14(12):e0226145. doi: 10.1371/journal.pone.0226145. eCollection 2019. PLoS One. 2019. PMID: 31809526 Free PMC article.
References
-
- Rosloniec EF, Cremer M, Kang AH, Myers LK, Brand DD. Collagen-induced arthritis. Curr Protoc Immunol. 2010;16:11–25. - PubMed
-
- Rosloniec EF, Brand DD, Myers LK, Whittington KB, Gumanovskaya M, Zaller DM, Woods A, Altmann DM, Stuart JM, Kang AH. An HLA-DR1 transgene confers susceptibility to collagen-induced arthritis elicited with human type II collagen. J Exp Med. 1997;16:1113–1122. doi: 10.1084/jem.185.6.1113. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
