

Pleiotropic effects of cancer cells' secreted factors on human stromal (mesenchymal) stem cells
- PMID: 24405819
- PMCID: PMC3854757
- DOI: 10.1186/scrt325
Pleiotropic effects of cancer cells' secreted factors on human stromal (mesenchymal) stem cells
Abstract
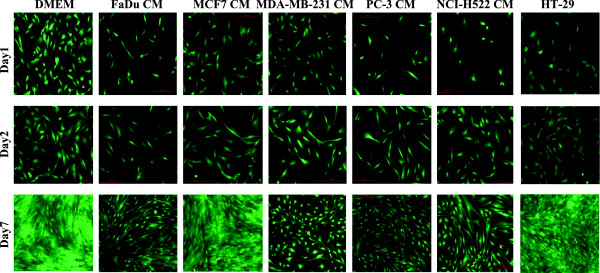
Introduction: Studying cancer tumors' microenvironment may reveal a novel role in driving cancer progression and metastasis. The biological interaction between stromal (mesenchymal) stem cells (MSCs) and cancer cells remains incompletely understood. Herein, we investigated the effects of tumor cells' secreted factors as represented by a panel of human cancer cell lines (breast (MCF7 and MDA-MB-231); prostate (PC-3); lung (NCI-H522); colon (HT-29) and head & neck (FaDu)) on the biological characteristics of MSCs.
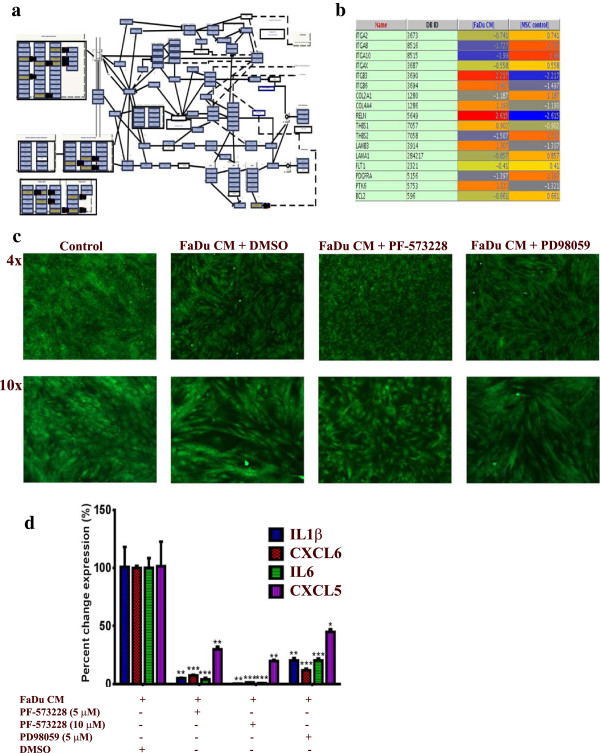
Methods: Morphological changes were assessed using fluorescence microscopy. Changes in gene expression were assessed using Agilent microarray and qRT-PCR. GeneSpring 12.1 and DAVID tools were used for bioinformatic and signaling pathway analyses. Cell migration was assessed using a transwell migration system. SB-431542, PF-573228 and PD98059 were used to inhibit transforming growth factor β (TGFβ), focal adhesion kinase (FAK), and mitogen activated protein kinase kinase (MAPKK) pathways, respectively. Interleukin-1β (IL1β) was measured using ELISA.
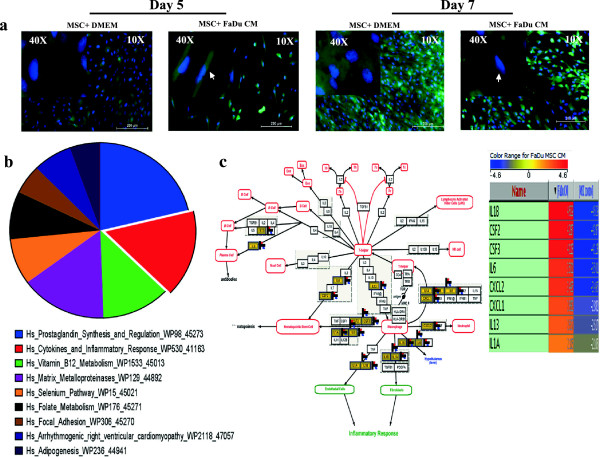
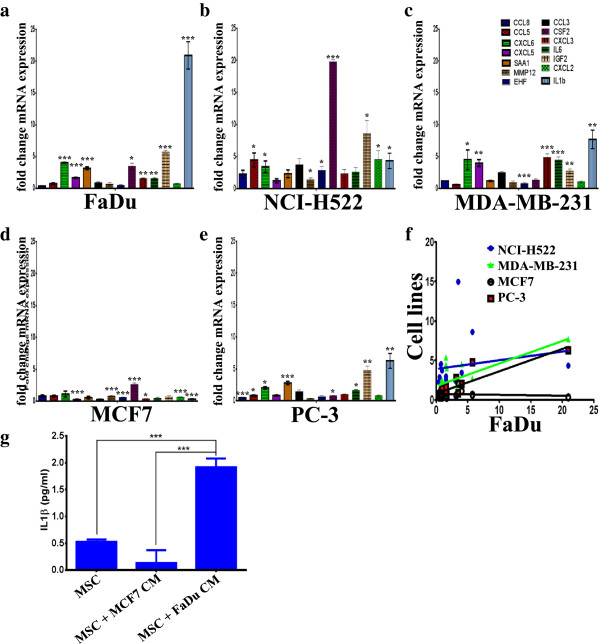
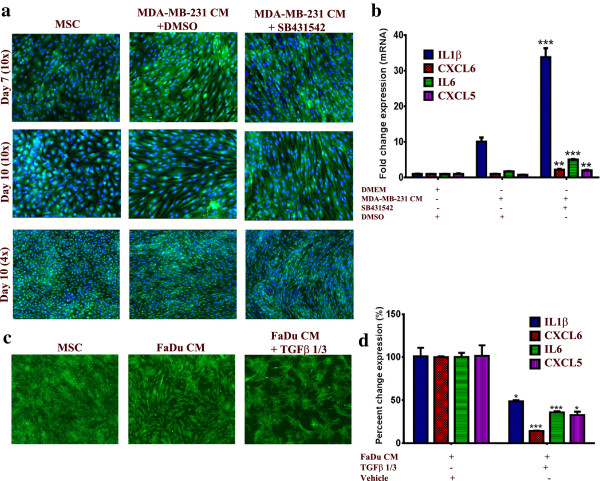
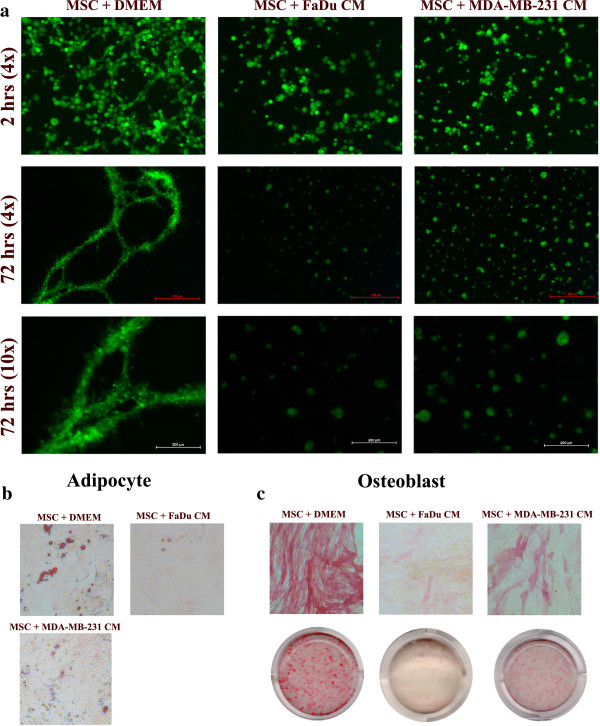
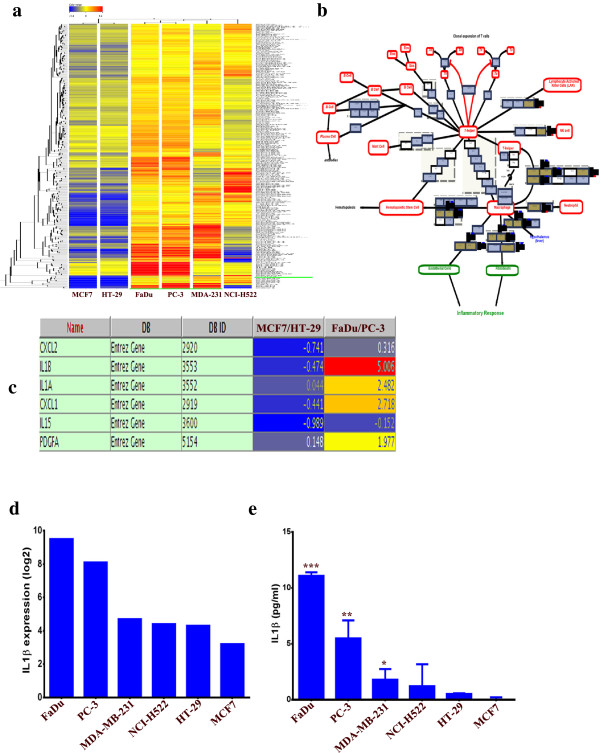
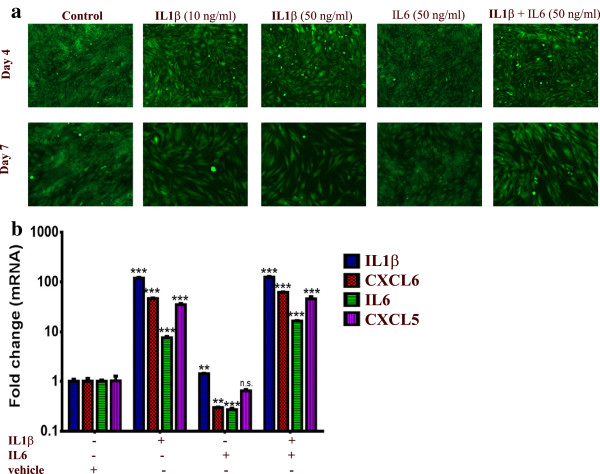
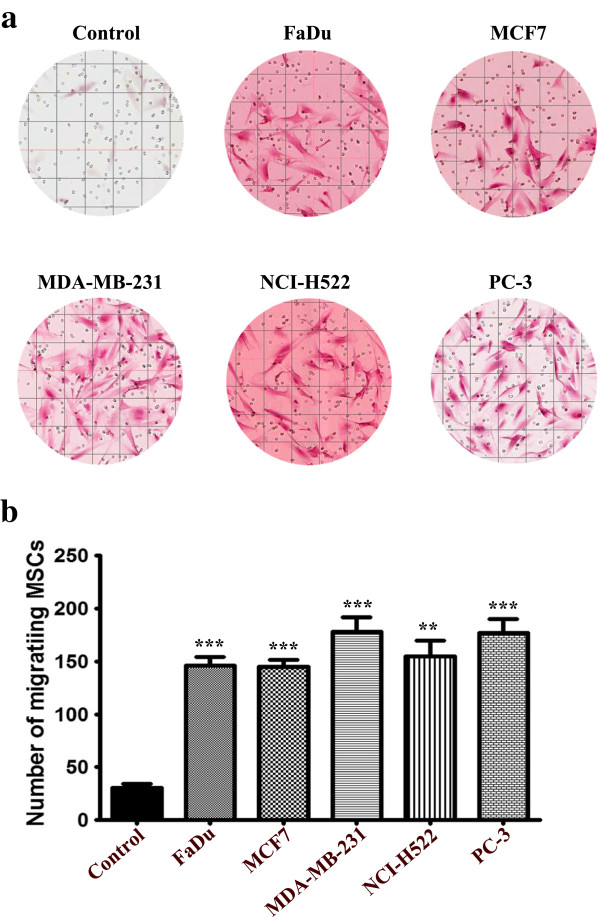
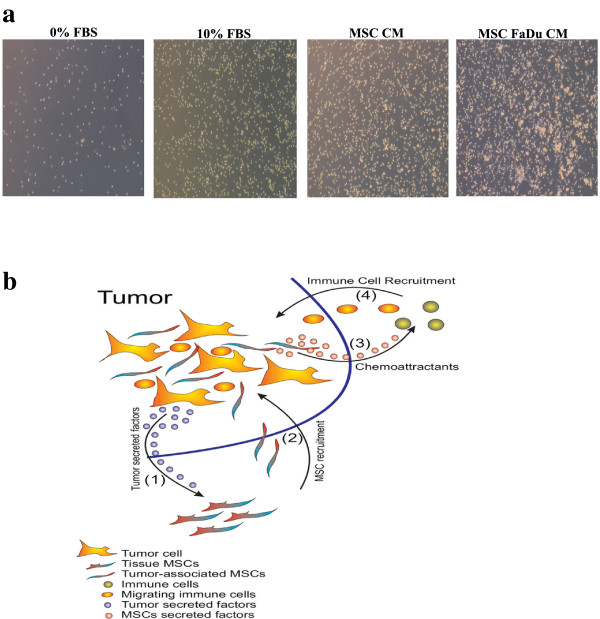
Results: MSCs exposed to secreted factors present in conditioned media (CM) from FaDu, MDA-MB-231, PC-3 and NCI-H522, but not from MCF7 and HT-29, developed an elongated, spindle-shaped morphology with bipolar processes. In association with phenotypic changes, genome-wide gene expression and bioinformatics analysis revealed an enhanced pro-inflammatory response of those MSCs. Pharmacological inhibitions of FAK and MAPKK severely impaired the pro-inflammatory response of MSCs to tumor CM (approximately 80% to 99%, and 55% to 88% inhibition, respectively), while inhibition of the TGFβ pathway was found to promote the pro-inflammatory response (approximately 3-fold increase). In addition, bioinformatics and pathway analysis of gene expression data from tumor cell lines combined with experimental validation revealed tumor-derived IL1β as one mediator of the pro-inflammatory phenotype observed in MSCs exposed to tumor CM.
Conclusions: Our data revealed tumor-derived IL1β as one mediator of the pro-inflammatory response in MSCs exposed to tumor CM, which was found to be positively regulated by FAK and MAPK signaling and negatively regulated by TGFβ signaling. Thus, our data support a model where MSCs could promote cancer progression through becoming pro-inflammatory cells within the cancer stroma.
Figures
Comment in
-
Biological effects of cancer-secreted factors on human mesenchymal stem cells.Stem Cell Res Ther. 2013 Nov 15;4(6):138. doi: 10.1186/scrt349. Stem Cell Res Ther. 2013. PMID: 24456712 Free PMC article.
References
-
- Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop D, Horwitz E. Minimal criteria for defining multipotent mesenchymal stromal cells, The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–317. doi: 10.1080/14653240600855905. - DOI - PubMed
-
- Kristensen LP, Chen L, Nielsen MO, Qanie DW, Kratchmarova I, Kassem M, Andersen JS. Temporal profiling and pulsed SILAC labeling identify novel secreted proteins during ex vivo osteoblast differentiation of human stromal stem cells. Mol Cell Proteomics. 2012;11:989–1007. doi: 10.1074/mcp.M111.012138. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
