

Analysis of inter-subunit contacts reveals the structural malleability of extracellular domains in platelet glycoprotein Ib-IX complex
- PMID: 24406065
- PMCID: PMC4137403
- DOI: 10.1111/jth.12437
Analysis of inter-subunit contacts reveals the structural malleability of extracellular domains in platelet glycoprotein Ib-IX complex
Abstract
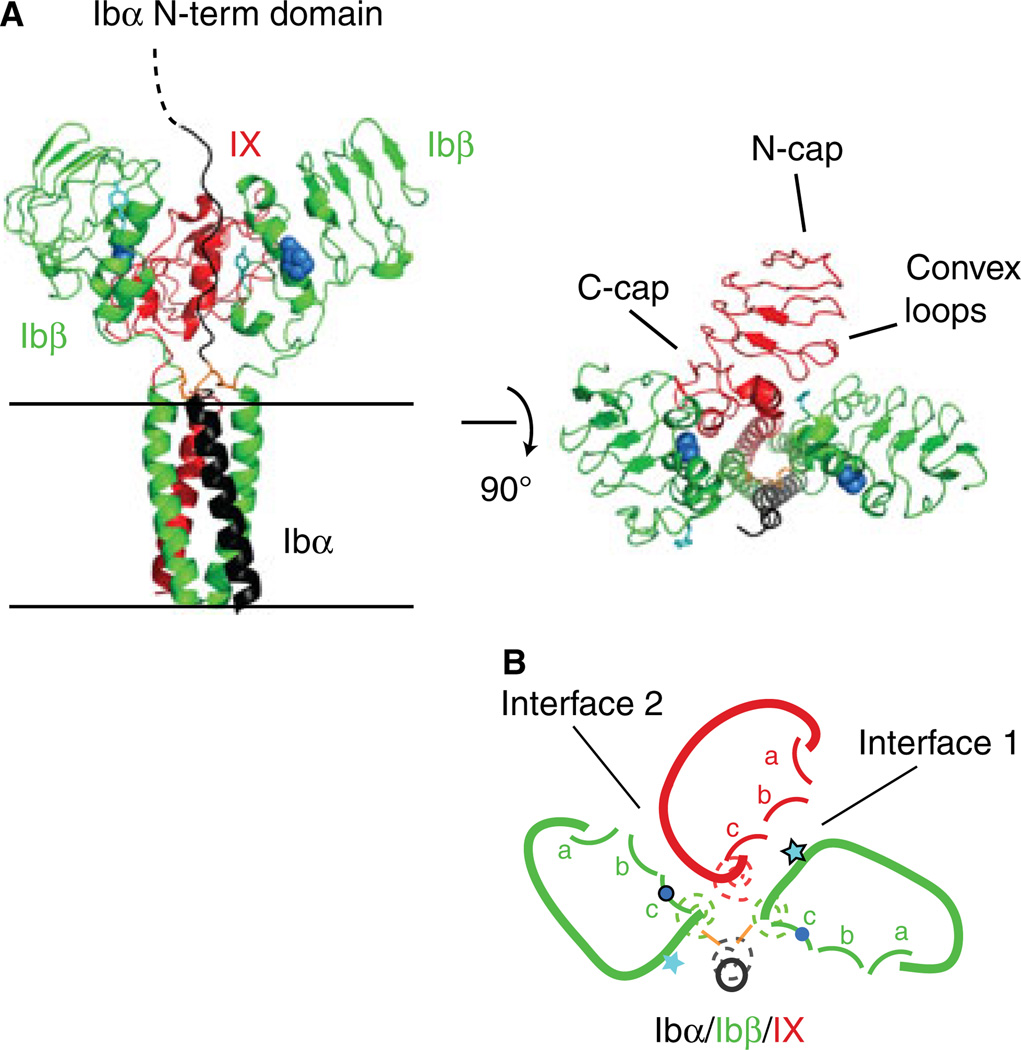
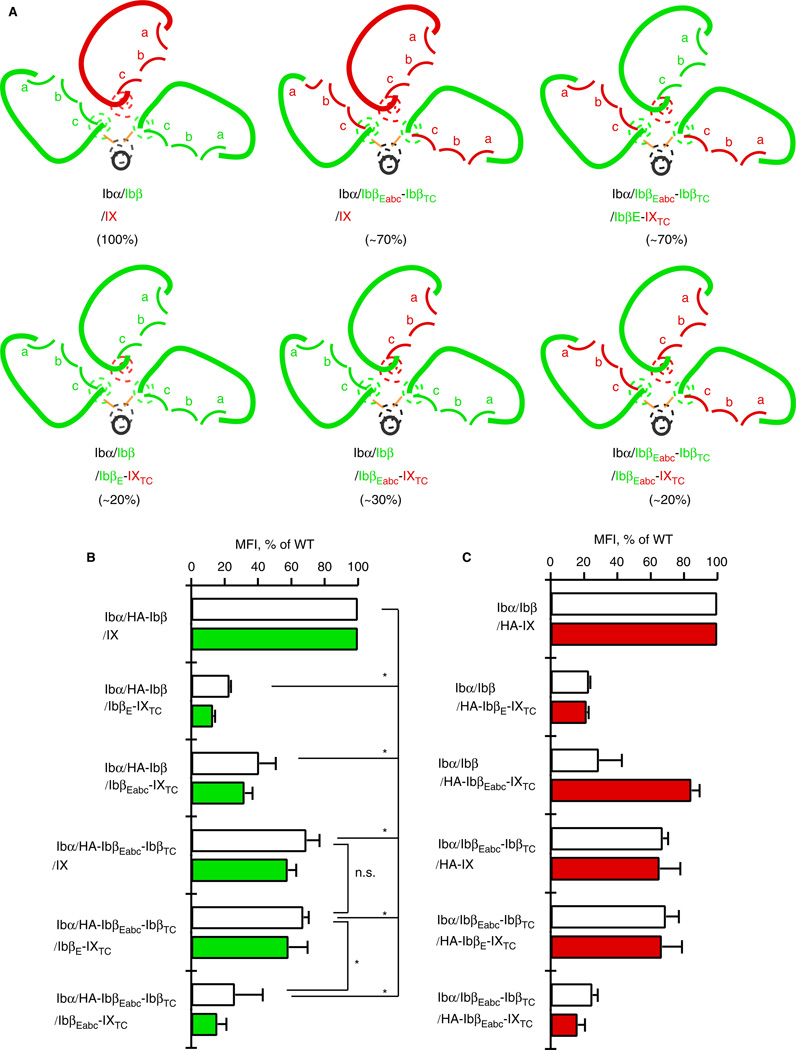
Background: The glycoprotein (GP)Ib-IX complex is critical to hemostasis and thrombosis. Its proper assembly is closely correlated with its surface expression level and requires cooperative interactions among extracellular and transmembrane domains of Ibα, Ibβ and IX subunits. Two interfaces have been previously identified between the extracellular domains of Ibβ and IX.
Objective: To understand how extracellular domains interact in GPIb-IX.
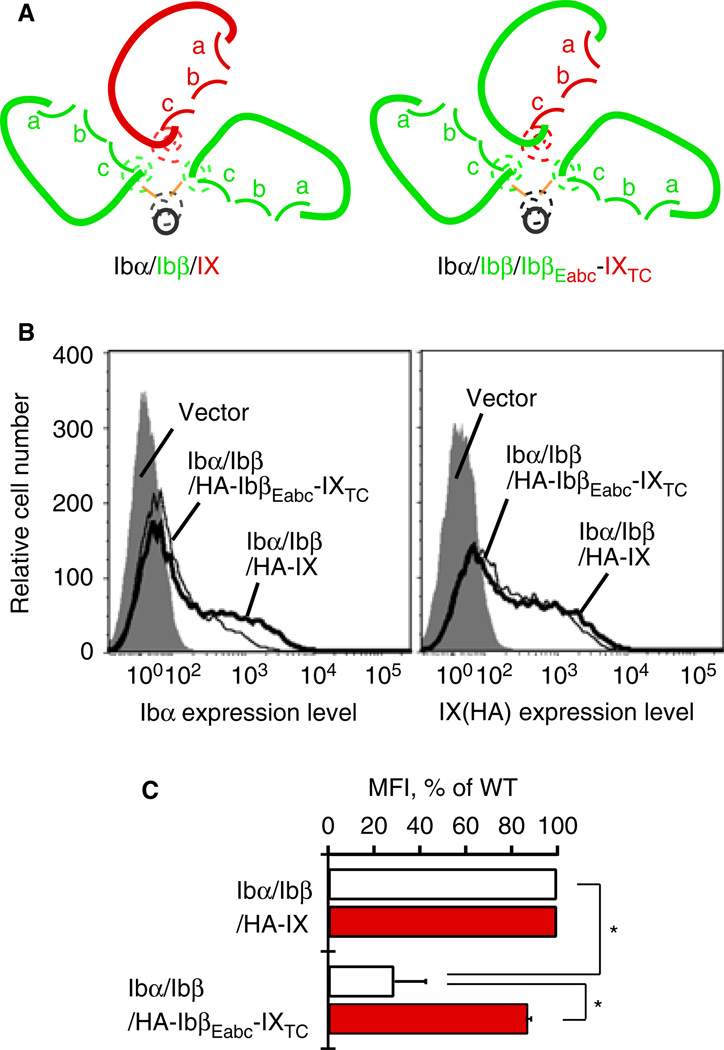
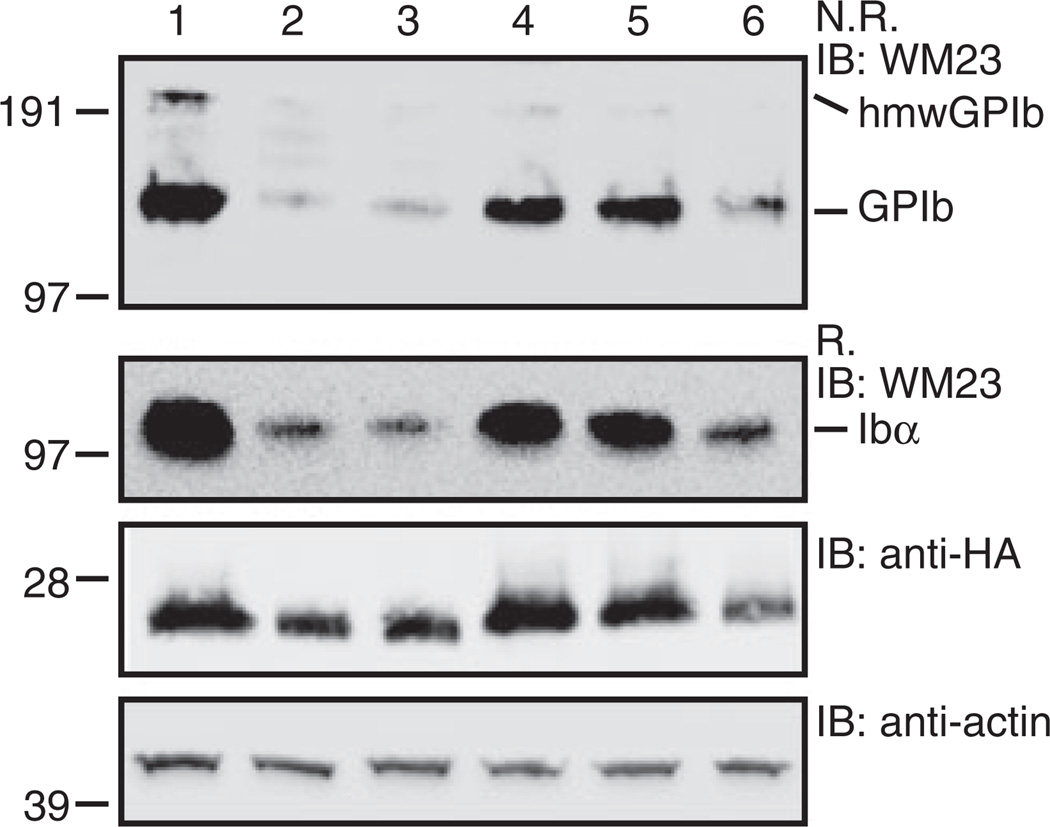
Methods: The Ibβ extracellular domain (IbβE ) or the IX counterpart (IXE ) in GPIb-IX was replaced with a well-folded IbβE /IXE chimera called IbβEabc , and the effect of domain replacement on assembly and expression of the receptor complex in transiently transfected Chinese hamster ovary cells was analyzed.
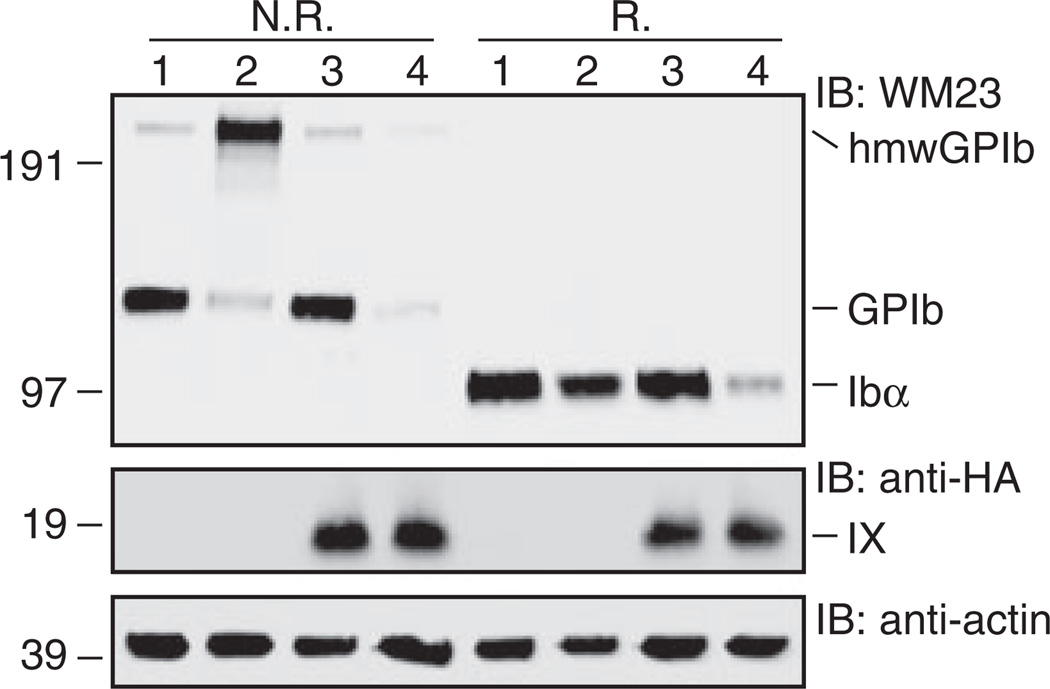
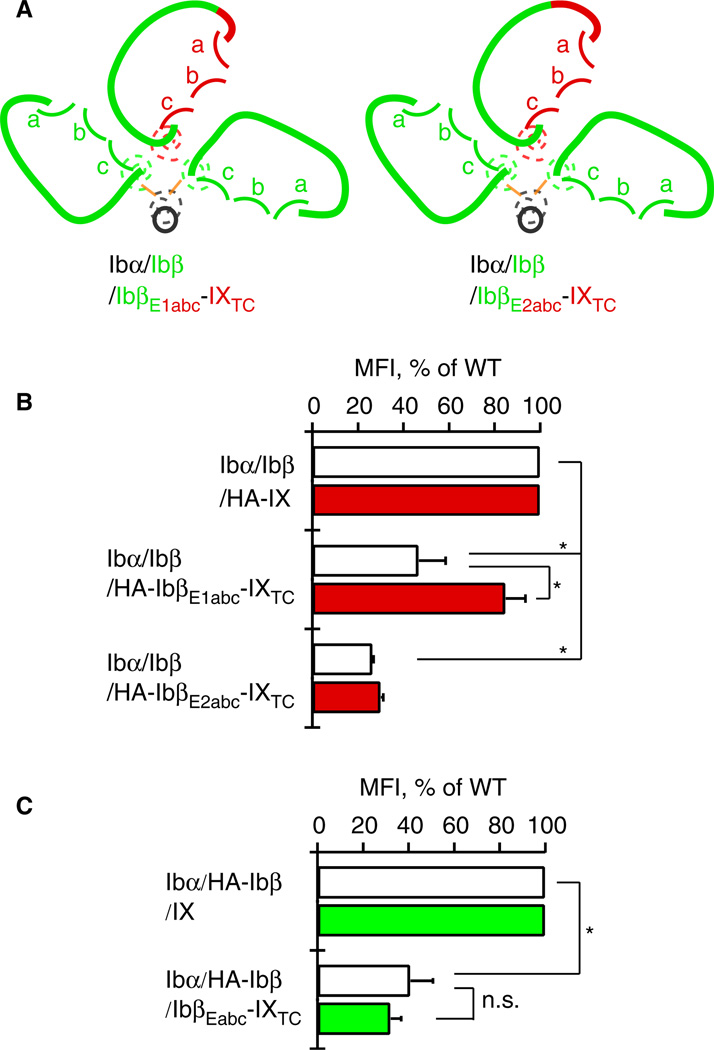
Results: Replacing IXE with IbβEabc in GPIb-IX retained interface 1 but not interface 2 between the extracellular domains. While this domain replacement preserved complex integrity, the expression levels of Ibβ and Ibα were significantly reduced. Additional domain replacement with IbβEabc or IbβE in GPIb-IX produced the complex at disparate expression levels that cannot be simply explained by two separate interfaces. In particular, when IbβE in GPIb-IX was replaced by IbβEabc , Ibα and IX were expressed at approximately 70% of the wild-type level. Their levels were not reduced when IXE was changed further to IbβE .
Conclusions: Our results demonstrate the importance of the association between Ibβ and IX extracellular domains for complex assembly and efficient expression, and provide evidence for the structural malleability of these domains that may accommodate and propagate conformational changes therein.
Keywords: Bernard-Soulier syndrome; leucine-rich repeat proteins; platelet glycoprotein GPIb-IX complex; protein-protein interaction domains; von Willebrand factor receptors.
© 2013 International Society on Thrombosis and Haemostasis.
Conflict of interest statement
The authors state that they have no conflict of interests.
Figures
References
-
- Berndt MC, Gregory C, Kabral A, Zola H, Fournier D, Castaldi PA. Purification and preliminary characterization of the glycoprotein Ib complex in the human platelet membrane. Eur J Biochem. 1985;151:637–649. - PubMed
-
- Huizinga EG, Tsuji S, Romijn RA, Schiphorst ME, de Groot PG, Sixma JJ, Gros P. Structures of glycoprotein Ibα and its complex with von Willebrand factor A1 domain. Science. 2002;297:1176–1179. - PubMed
-
- Dumas JJ, Kumar R, Seehra J, Somers WS, Mosyak L. Crystal structure of the GpIbα-thrombin complex essential for platelet aggregation. Science. 2003;301:222–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
