

Rab1a regulates sorting of early endocytic vesicles
- PMID: 24407591
- PMCID: PMC3949023
- DOI: 10.1152/ajpgi.00118.2013
Rab1a regulates sorting of early endocytic vesicles
Abstract
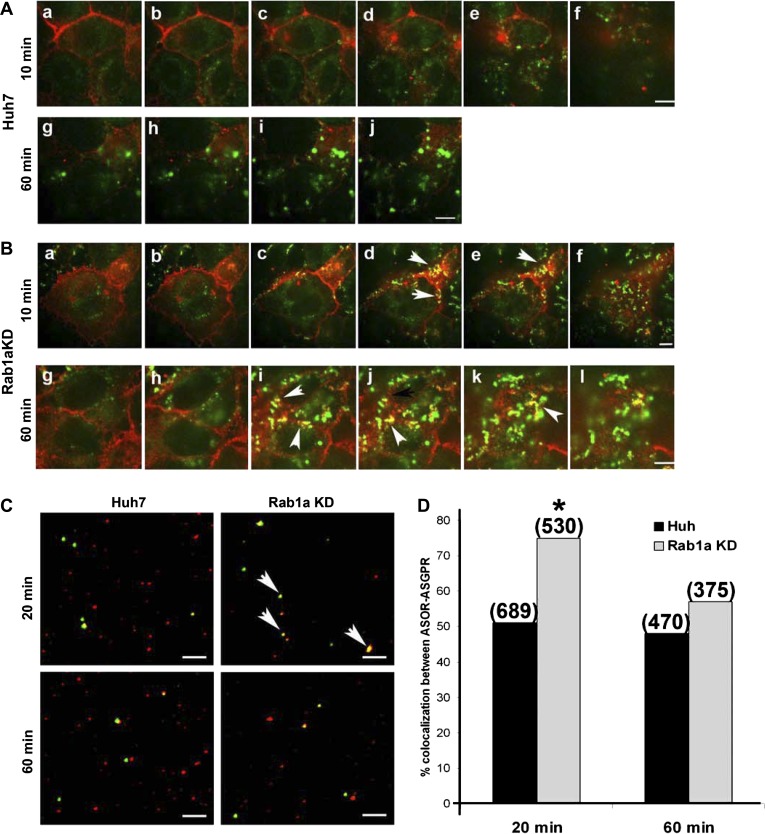
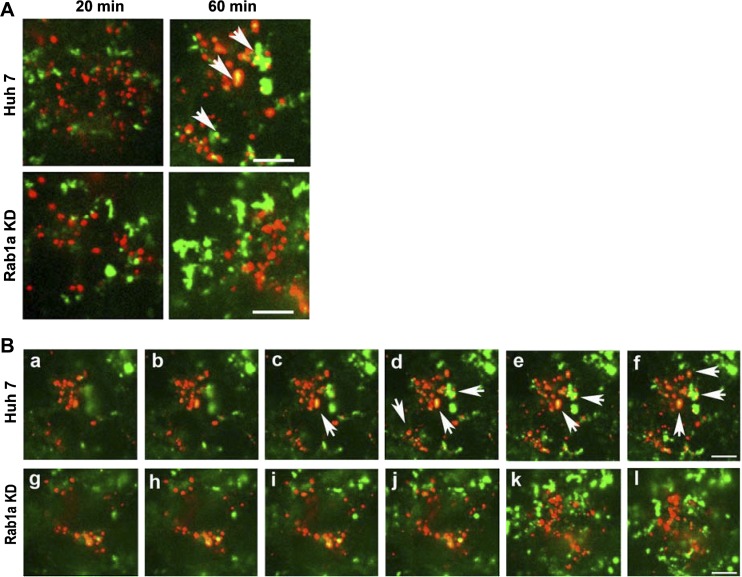
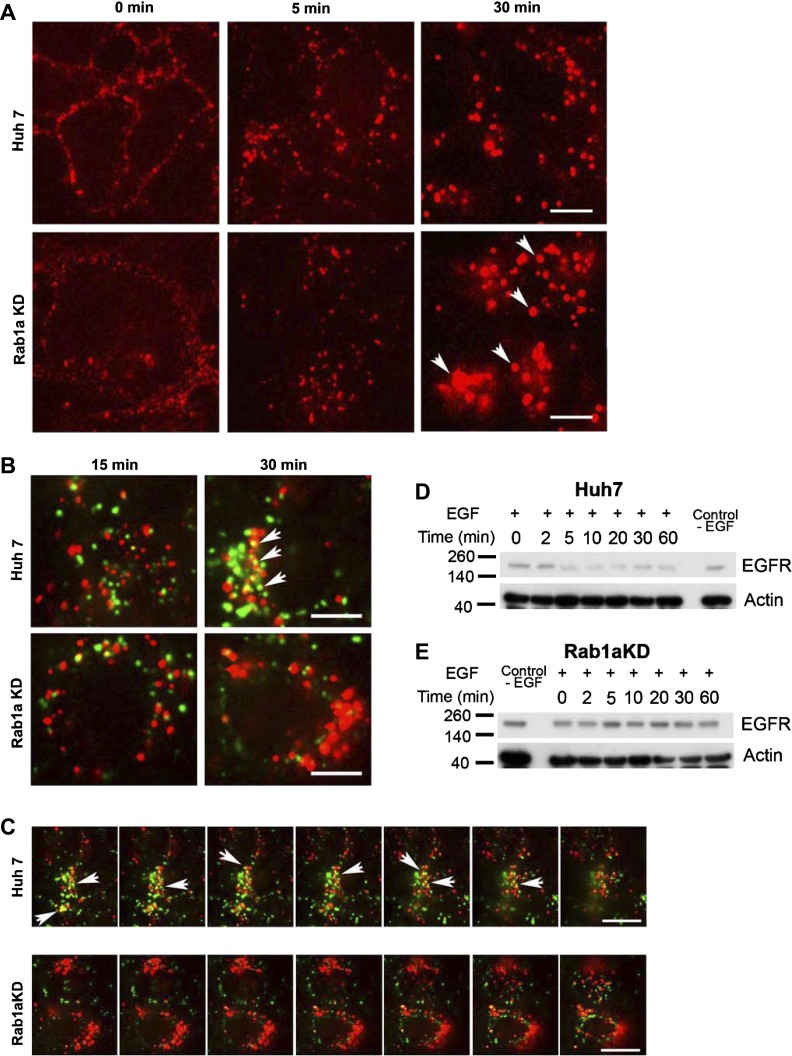
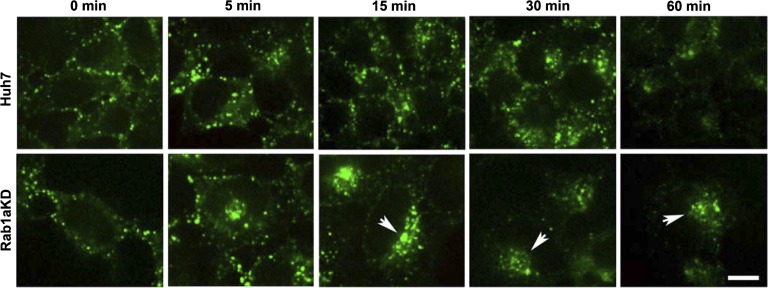
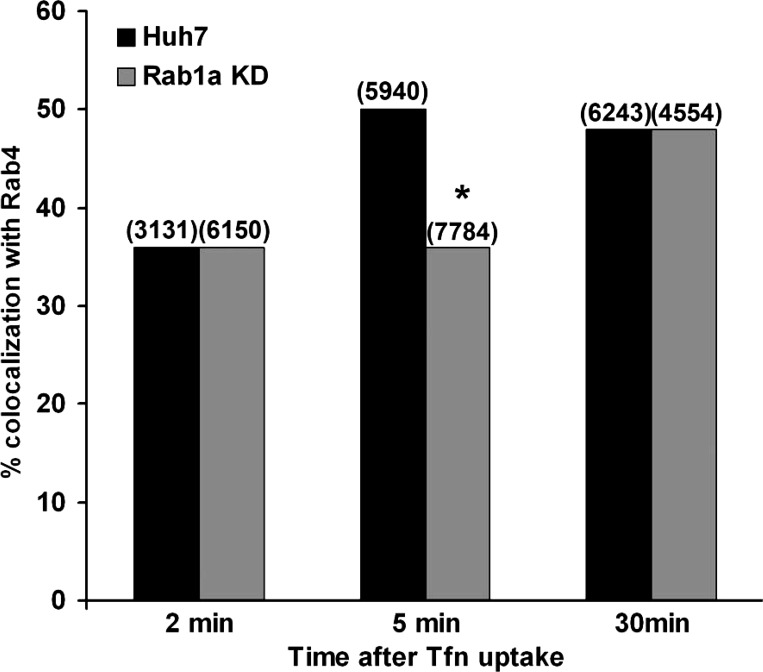
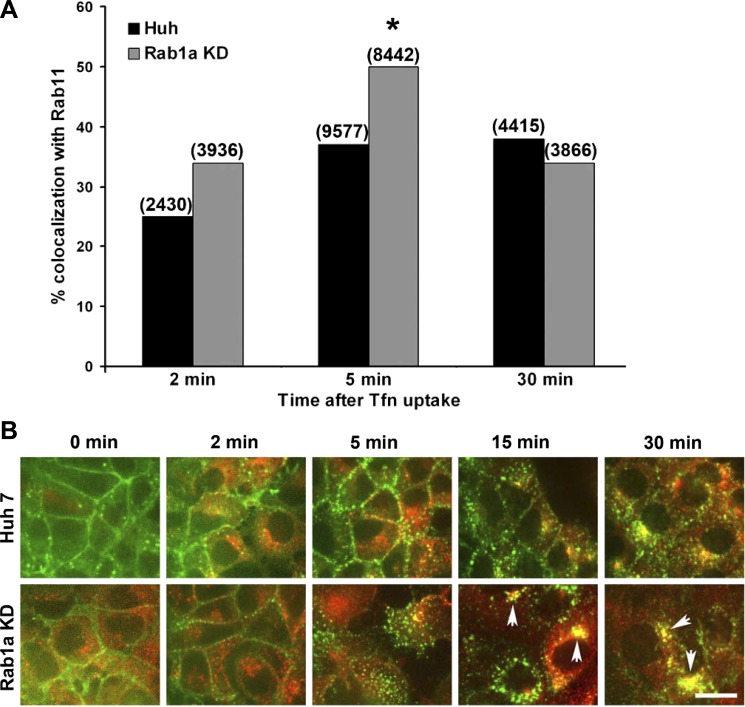
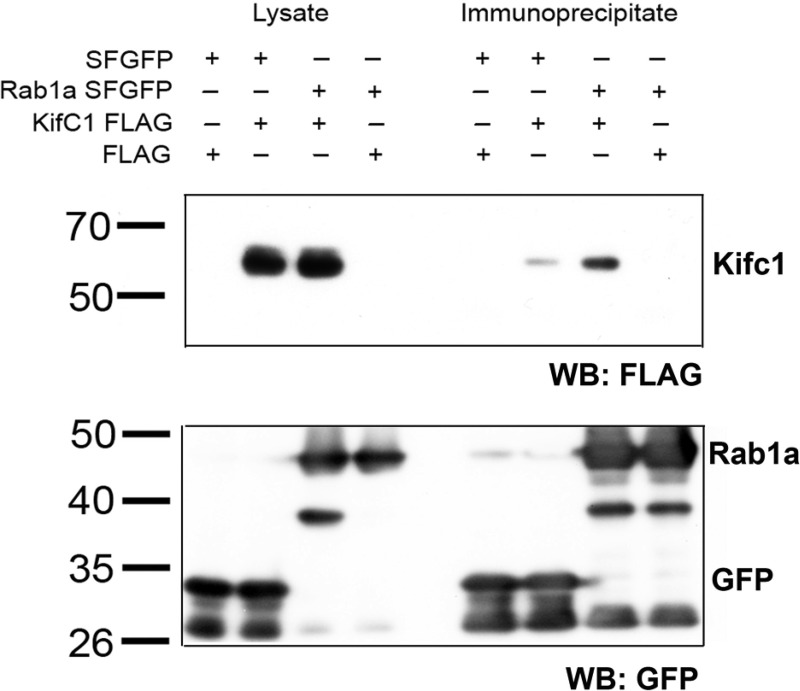
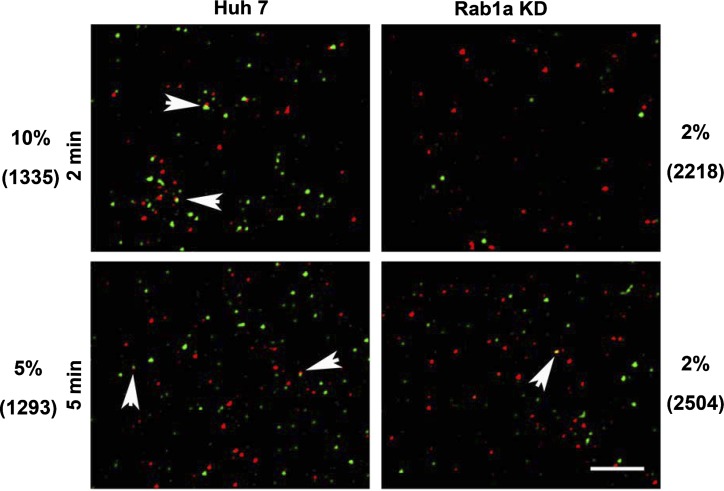
We previously reported that Rab1a is associated with asialoorosomucoid (ASOR)-containing early endocytic vesicles, where it is required for their microtubule-based motility. In Rab1a knockdown (KD) cell lines, ASOR failed to segregate from its receptor and, consequently, did not reach lysosomes for degradation, indicating a defect in early endosome sorting. Although Rab1 is required for Golgi/endoplasmic reticulum trafficking, this process was unaffected, likely due to retained expression of Rab1b in these cells. The present study shows that Rab1a has a more general role in endocytic vesicle processing that extends to EGF and transferrin (Tfn) trafficking. Compared with results in control Huh7 cells, EGF accumulated in aggregates within Rab1a KD cells, failing to reach lysosomal compartments. Tfn, a prototypical example of recycling cargo, accumulated in a Rab11-mediated slow-recycling compartment in Rab1a KD cells, in contrast to control cells, which sort Tfn into a fast-recycling Rab4 compartment. These data indicate that Rab1a is an important regulator of early endosome sorting for multiple cargo species. The effectors and accessory proteins recruited by Rab1a to early endocytic vesicles include the minus-end-directed kinesin motor KifC1, while others remain to be discovered.
Keywords: EGF; Rab1a; endocytosis; sorting; transferrin.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources