

The regenerative potential of parietal epithelial cells in adult mice
- PMID: 24408873
- PMCID: PMC3968497
- DOI: 10.1681/ASN.2013050481
The regenerative potential of parietal epithelial cells in adult mice
Abstract
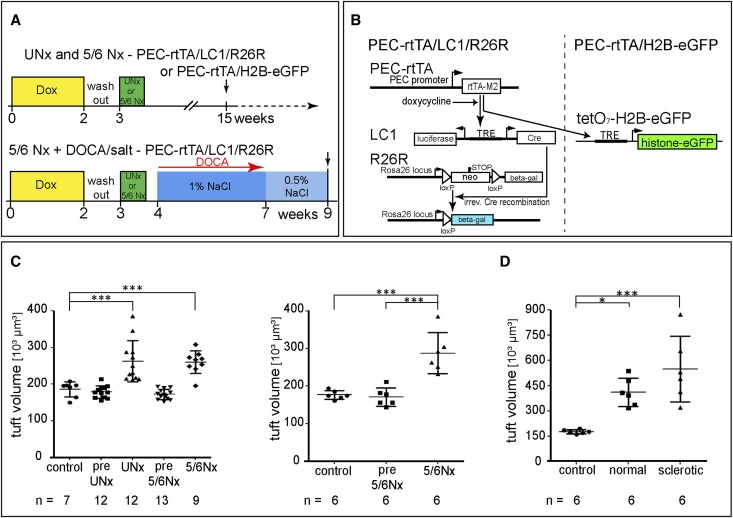
Previously, we showed that some podocytes in juvenile mice are recruited from cells lining Bowman's capsule, suggesting that parietal epithelial cells (PECs) are a progenitor cell population for podocytes. To investigate whether PECs also replenish podocytes in adult mice, PECs were genetically labeled in an irreversible fashion in 5-week-old mice. No significant increase in labeled podocytes was observed, even after 18 months. To accelerate a potential regenerative mechanism, progressive glomerular hypertrophy was induced by progressive partial nephrectomies. Again, no significant podocyte replenishment was observed. Rather, labeled PECs exclusively invaded segments of the tuft affected by glomerulosclerosis, consistent with our previous findings. We next reassessed PEC recruitment in juvenile mice using a different reporter mouse and confirmed significant recruitment of labeled PECs onto the glomerular tuft. Moreover, some labeled cells on Bowman's capsule expressed podocyte markers, and cells on Bowman's capsule were also directly labeled in juvenile podocyte-specific Pod-rtTA transgenic mice. In 6-week-old mice, however, cells on Bowman's capsule no longer expressed podocyte-specific markers. Similarly, in human kidneys, some cells on Bowman's capsule expressed the podocyte marker synaptopodin from 2 weeks to 2 years of age but not at 7 years of age. In summary, podocyte regeneration from PECs could not be detected in aging mice or models of glomerular hypertrophy. We propose that a small fraction of committed podocytes reside on Bowman's capsule close to the vascular stalk and are recruited onto the glomerular tuft during infancy to adolescence in mice and humans.
Figures








References
-
- Kriz W, Gretz N, Lemley KV: Progression of glomerular diseases: Is the podocyte the culprit? Kidney Int 54: 687–697, 1998 - PubMed
-
- Wharram BL, Goyal M, Wiggins JE, Sanden SK, Hussain S, Filipiak WE, Saunders TL, Dysko RC, Kohno K, Holzman LB, Wiggins RC: Podocyte depletion causes glomerulosclerosis: Diphtheria toxin-induced podocyte depletion in rats expressing human diphtheria toxin receptor transgene. J Am Soc Nephrol 16: 2941–2952, 2005 - PubMed
-
- Wiggins RC: The spectrum of podocytopathies: A unifying view of glomerular diseases. Kidney Int 71: 1205–1214, 2007 - PubMed
-
- Nagata M, Yamaguchi Y, Komatsu Y, Ito K: Mitosis and the presence of binucleate cells among glomerular podocytes in diseased human kidneys. Nephron 70: 68–71, 1995 - PubMed
-
- Pabst R, Sterzel RB: Cell renewal of glomerular cell types in normal rats. An autoradiographic analysis. Kidney Int 24: 626–631, 1983 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
