

Key role for CD4 T cells during mixed antibody-mediated rejection of renal allografts
- PMID: 24410909
- PMCID: PMC4128005
- DOI: 10.1111/ajt.12596
Key role for CD4 T cells during mixed antibody-mediated rejection of renal allografts
Abstract
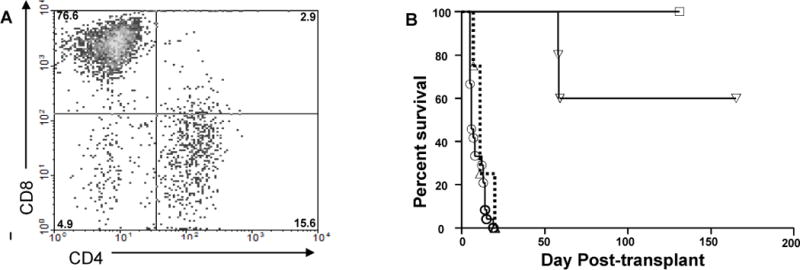
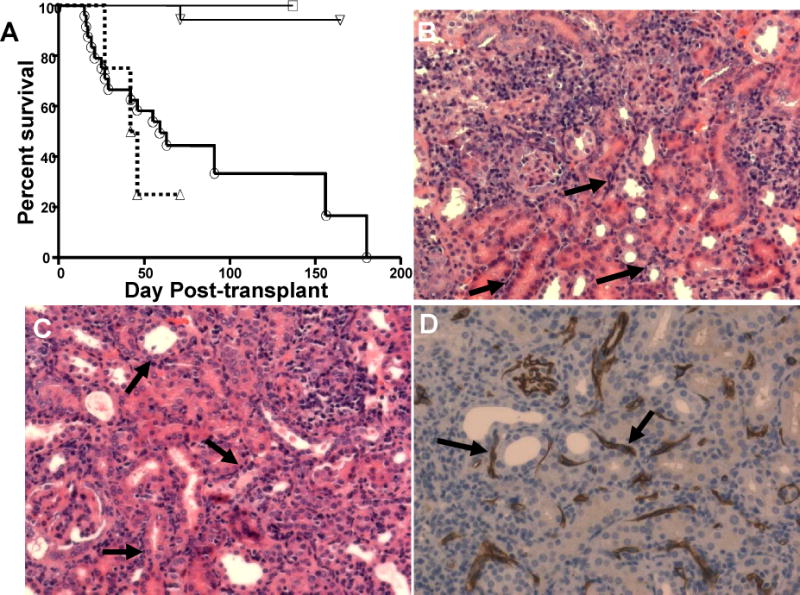
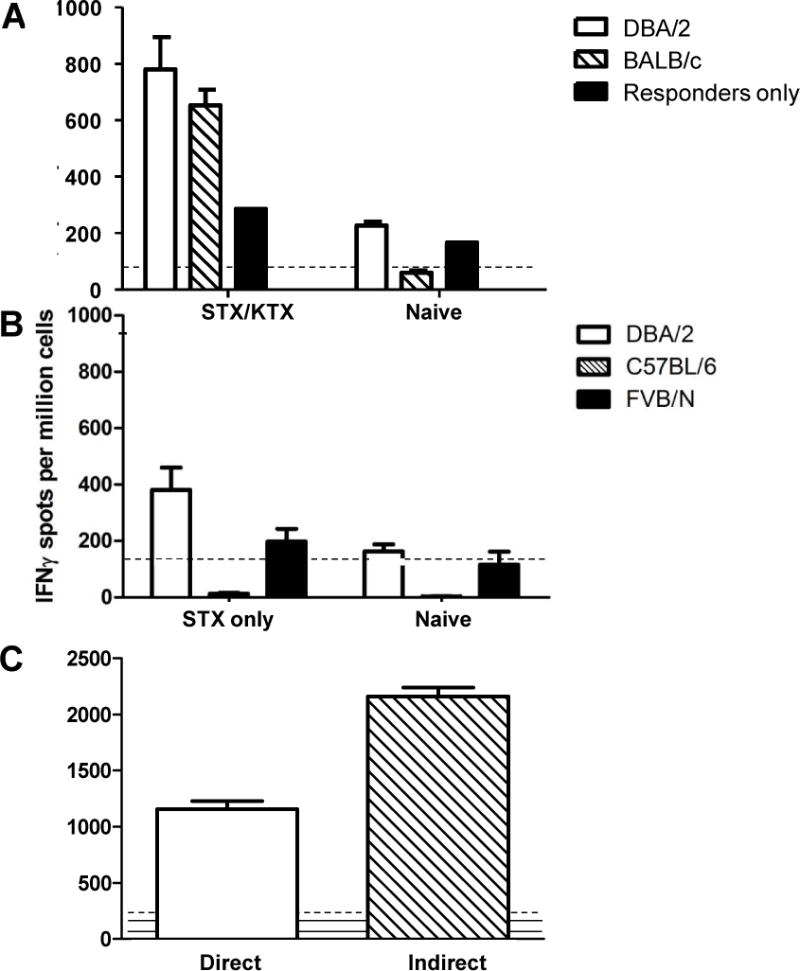
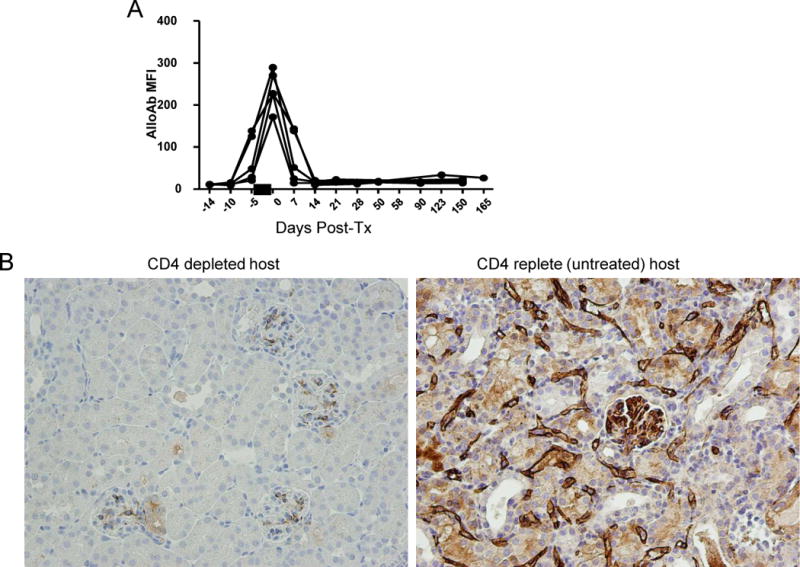
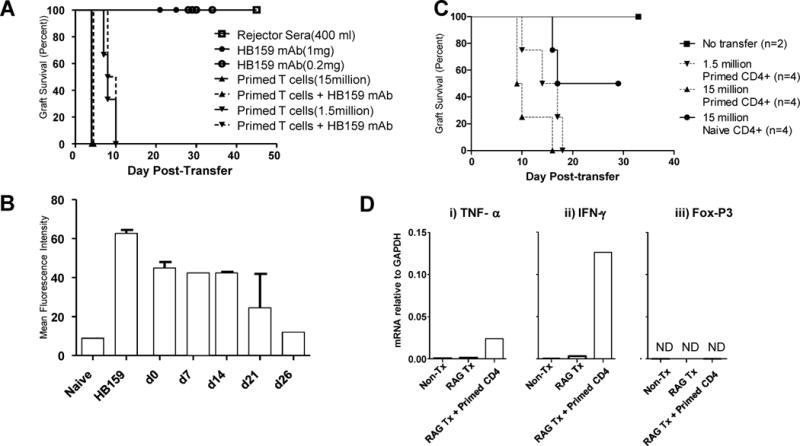
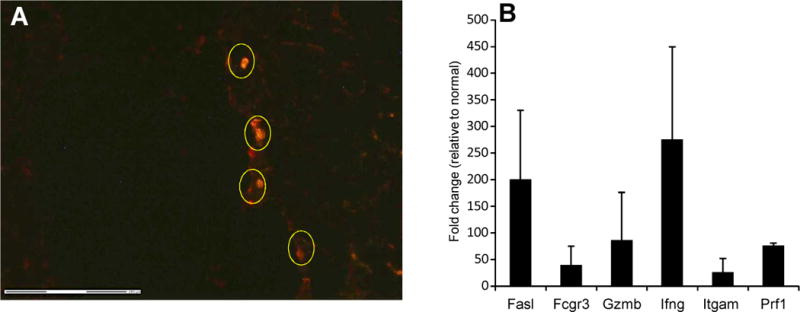
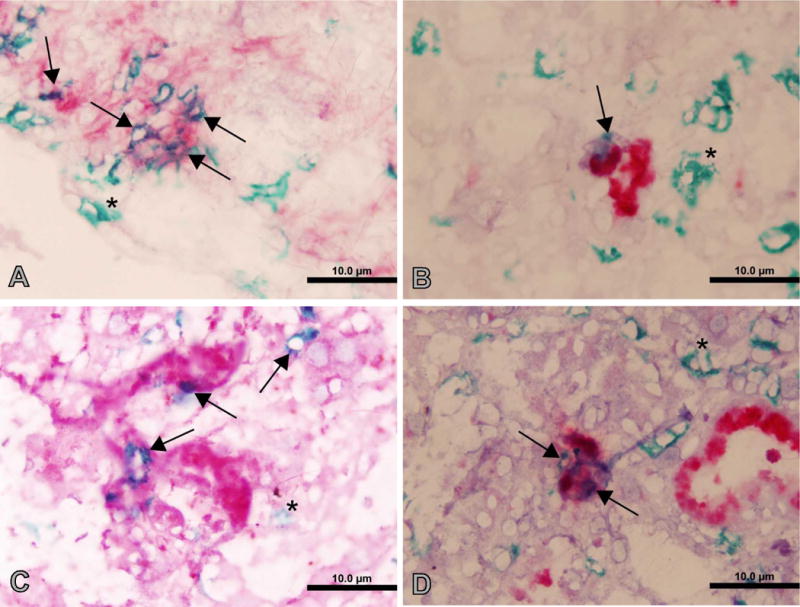
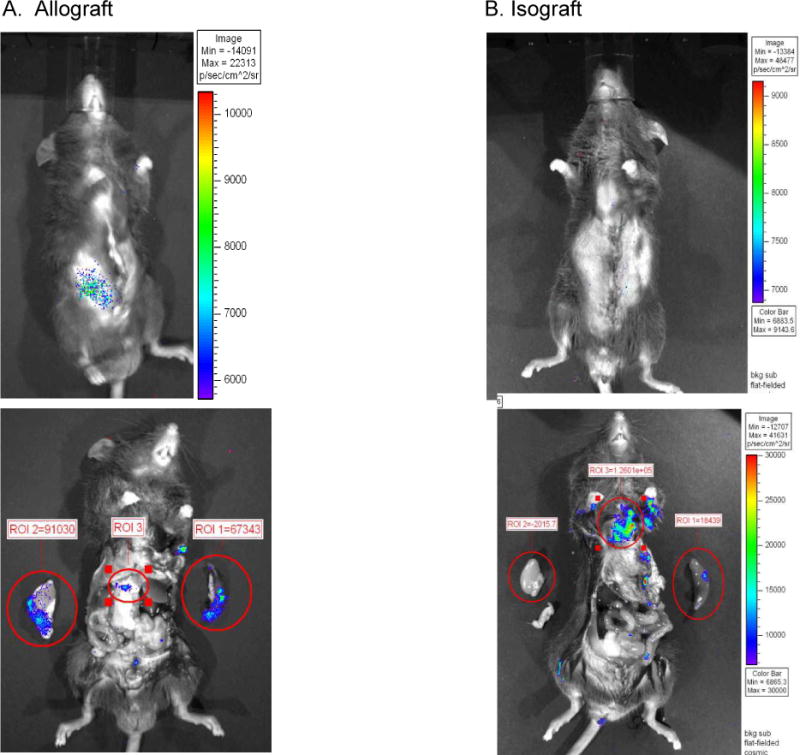
We utilized mouse models to elucidate the immunologic mechanisms of functional graft loss during mixed antibody-mediated rejection of renal allografts (mixed AMR), in which humoral and cellular responses to the graft occur concomitantly. Although the majority of T cells in the graft at the time of rejection were CD8 T cells with only a minor population of CD4 T cells, depletion of CD4 but not CD8 cells prevented acute graft loss during mixed AMR. CD4 depletion eliminated antidonor alloantibodies and conferred protection from destruction of renal allografts. ELISPOT revealed that CD4 T effectors responded to donor alloantigens by both the direct and indirect pathways of allorecognition. In transfer studies, CD4 T effectors primed to donor alloantigens were highly effective at promoting acute graft dysfunction, and exhibited the attributes of effector T cells. Laser capture microdissection and confirmatory immunostaining studies revealed that CD4 T cells infiltrating the graft produced effector molecules with graft destructive potential. Bioluminescent imaging confirmed that CD4 T effectors traffic to the graft site in immune replete hosts. These data document that host CD4 T cells can promote acute dysfunction of renal allografts by directly mediating graft injury in addition to facilitating antidonor alloantibody responses.
Keywords: Adoptive transfer; ELISPOT; T cell-mediated rejection; antibody-mediated rejection; graft infiltrating lymphocytes.
© Copyright 2014 The American Society of Transplantation and the American Society of Transplant Surgeons.
Conflict of interest statement
The authors of this manuscript have no conflicts of interest to disclose as described by the American Journal of Transplantation.
Figures
References
-
- Collins AB, Schneeberger EE, Pascual MA, Saidman SL, Williams WW, Tolkoff-Rubin N, et al. Complement activation in acute humoral renal allograft rejection: diagnostic significance of C4d deposits in peritubular capillaries. J Am Soc Nephrol. 1999 Oct;10(10):2208–14. Epub 1999/10/03. eng. - PubMed
-
- Bohmig GA, Exner M, Habicht A, Schillinger M, Lang U, Kletzmayr J, et al. Capillary C4d deposition in kidney allografts: a specific marker of alloantibody-dependent graft injury. J Am Soc Nephrol. 2002 Apr;13(4):1091–9. Epub 2002/03/26. eng. - PubMed
-
- Nakashima S, Qian Z, Rahimi S, Wasowska BA, Baldwin WM., 3rd Membrane attack complex contributes to destruction of vascular integrity in acute lung allograft rejection. J Immunol. 2002 Oct 15;169(8):4620–7. Epub 2002/10/09. eng. - PubMed
-
- Colvin RB. Antibody-mediated renal allograft rejection: diagnosis and pathogenesis. J Am Soc Nephrol. 2007 Apr;18(4):1046–56. Epub 2007/03/16. eng. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
