

Transient potential gradients and impedance measures of tethered bilayer lipid membranes: pore-forming peptide insertion and the effect of electroporation
- PMID: 24411250
- PMCID: PMC3907230
- DOI: 10.1016/j.bpj.2013.11.1121
Transient potential gradients and impedance measures of tethered bilayer lipid membranes: pore-forming peptide insertion and the effect of electroporation
Abstract
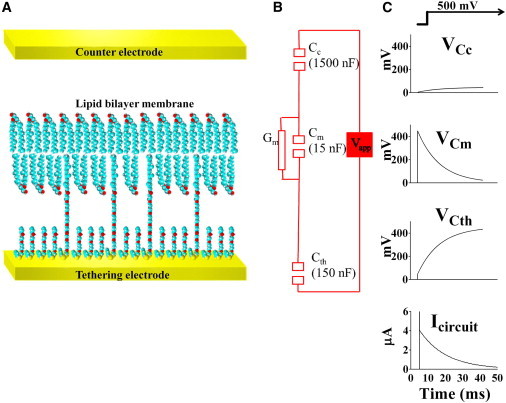
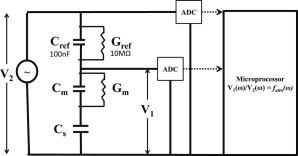
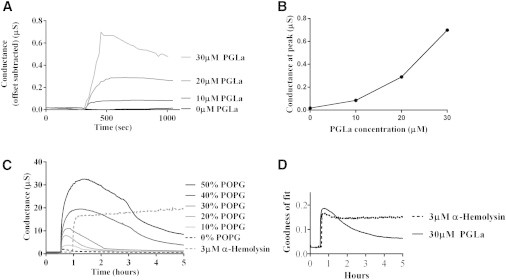
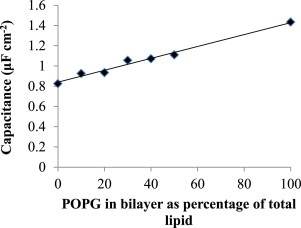
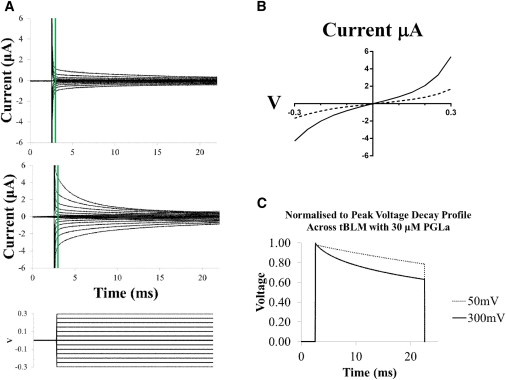
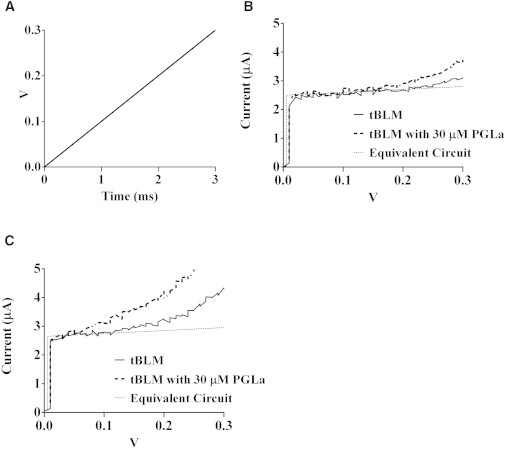
In this work, we present experimental data, supported by a quantitative model, on the generation and effect of potential gradients across a tethered bilayer lipid membrane (tBLM) with, to the best of our knowledge, novel architecture. A challenge to generating potential gradients across tBLMs arises from the tethering coordination chemistry requiring an inert metal such as gold, resulting in any externally applied voltage source being capacitively coupled to the tBLM. This in turn causes any potential across the tBLM assembly to decay to zero in milliseconds to seconds, depending on the level of membrane conductance. Transient voltages applied to tBLMs by pulsed or ramped direct-current amperometry can, however, provide current-voltage (I/V) data that may be used to measure the voltage dependency of the membrane conductance. We show that potential gradients >~150 mV induce membrane defects that permit the insertion of pore-forming peptides. Further, we report here the novel (to our knowledge) use of real-time modeling of conventional low-voltage alternating-current impedance spectroscopy to identify whether the conduction arising from the insertion of a polypeptide is uniform or heterogeneous on scales of nanometers to micrometers across the membrane. The utility of this tBLM architecture and these techniques is demonstrated by characterizing the resulting conduction properties of the antimicrobial peptide PGLa.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Heinrich F., Ng T., Lösche M. A new lipid anchor for sparsely tethered bilayer lipid membranes. Langmuir. 2009;25:4219–4229. - PubMed
-
- Cornell B.A. Memembrane-based biosensors. In: Ligler F., Taitt C.A.R., editors. Optical Biosensors: Present and Future. Elsevier Science; Amsterdam: 2002. pp. 457–459.
-
- Cornell B.A., Braach-Maksvytis V.L.B., Pace R.J. A biosensor that uses ion-channel switches. Nature. 1997;387:580–583. - PubMed
-
- Krishna G., Schulte J., Osman P.D. Tethered bilayer membranes containing ionic reservoirs: selectivity and conductance. Langmuir. 2003;19:2294–2305.
-
- Krishna G., Schulte J., Osman P.D. Tethered bilayer membranes containing ionic reservoirs: the interfacial capacitance. Langmuir. 2001;17:4858–4866.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
