

Proatherogenic conditions promote autoimmune T helper 17 cell responses in vivo
- PMID: 24412615
- PMCID: PMC3955722
- DOI: 10.1016/j.immuni.2013.11.021
Proatherogenic conditions promote autoimmune T helper 17 cell responses in vivo
Abstract
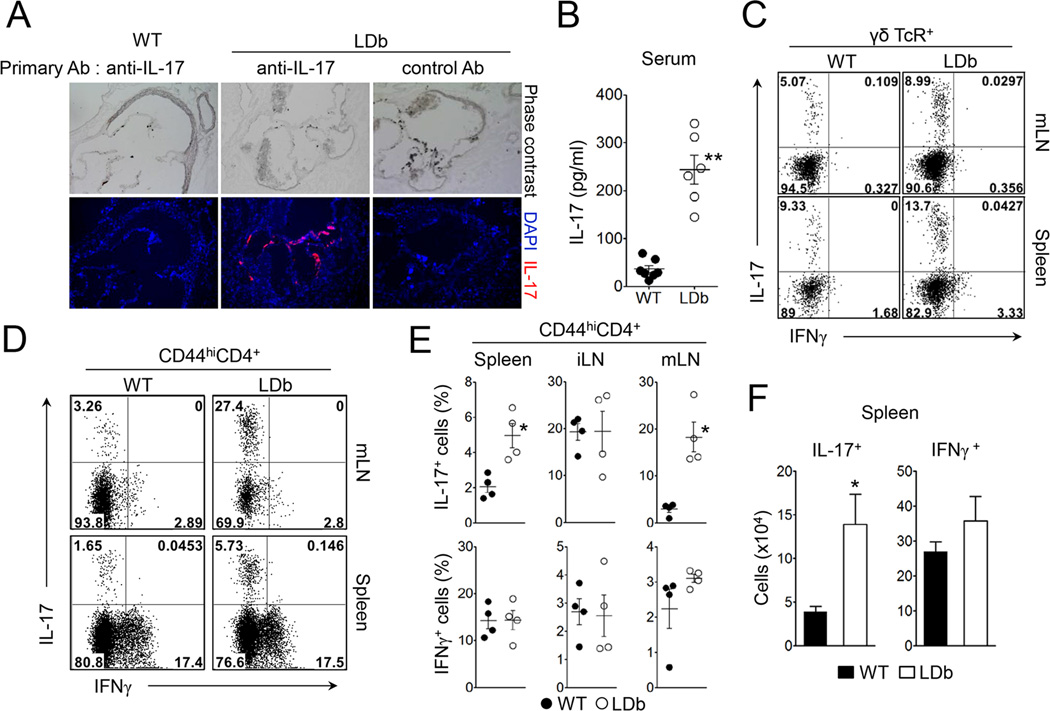
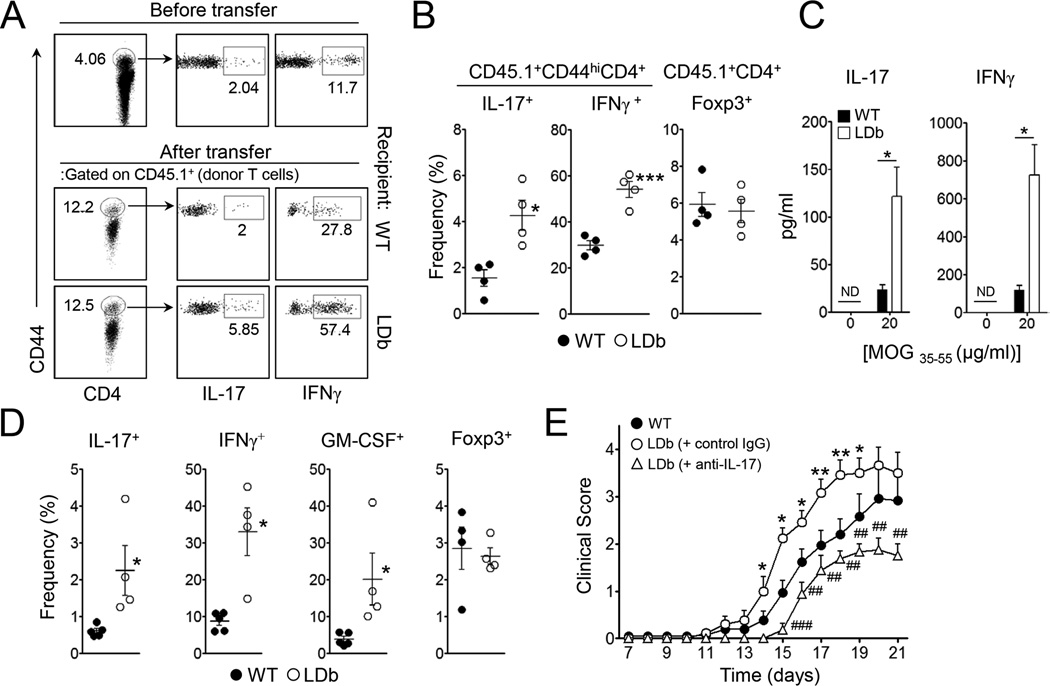
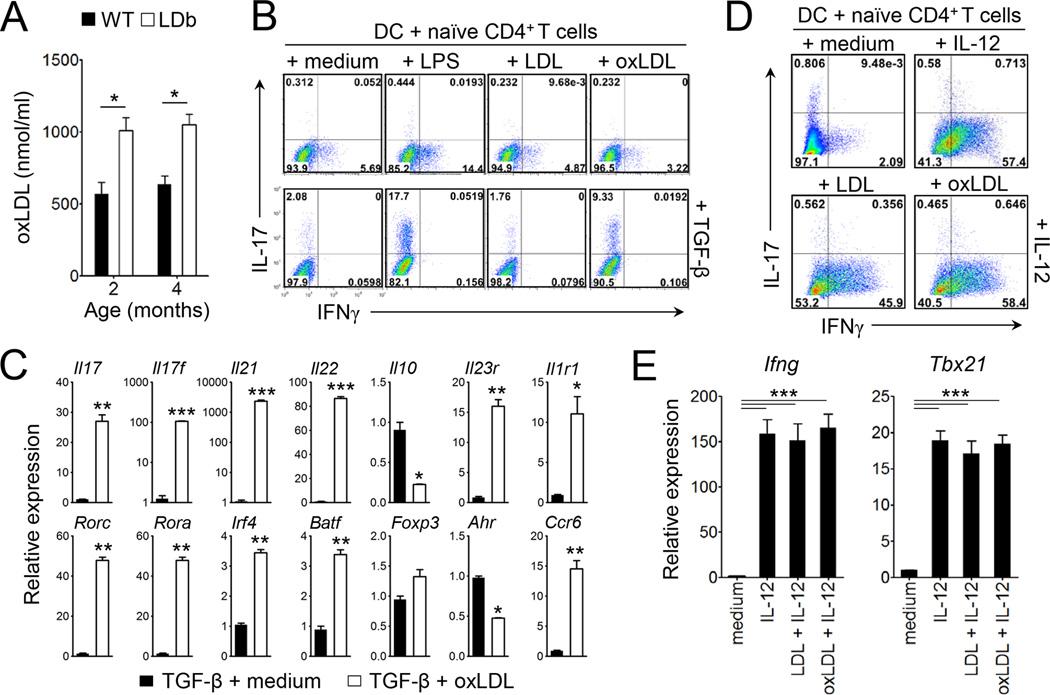
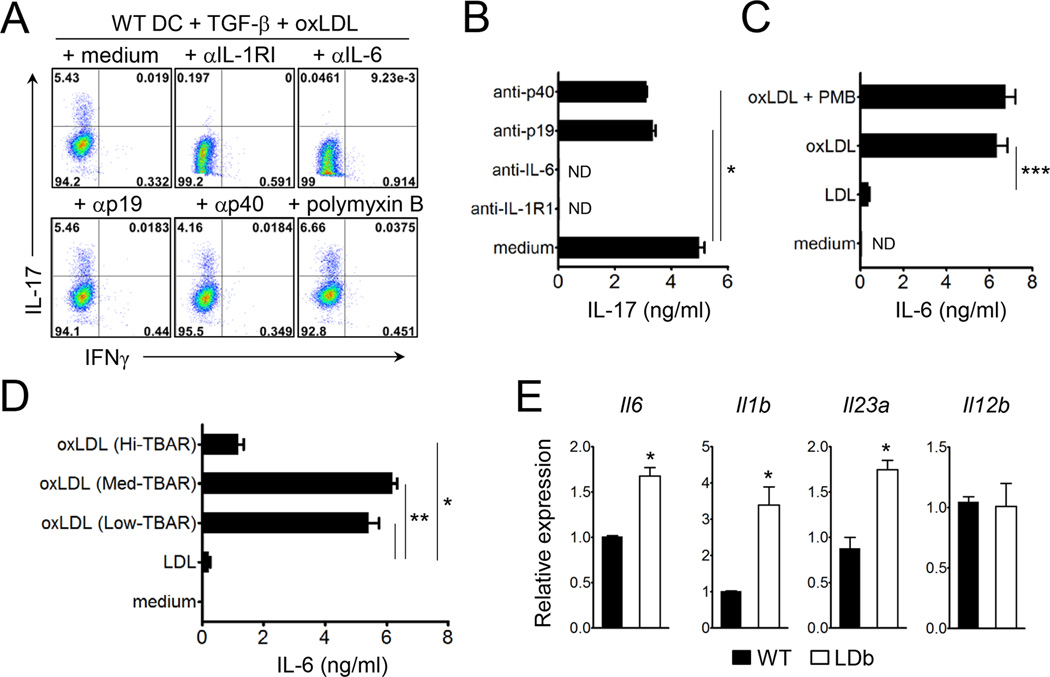
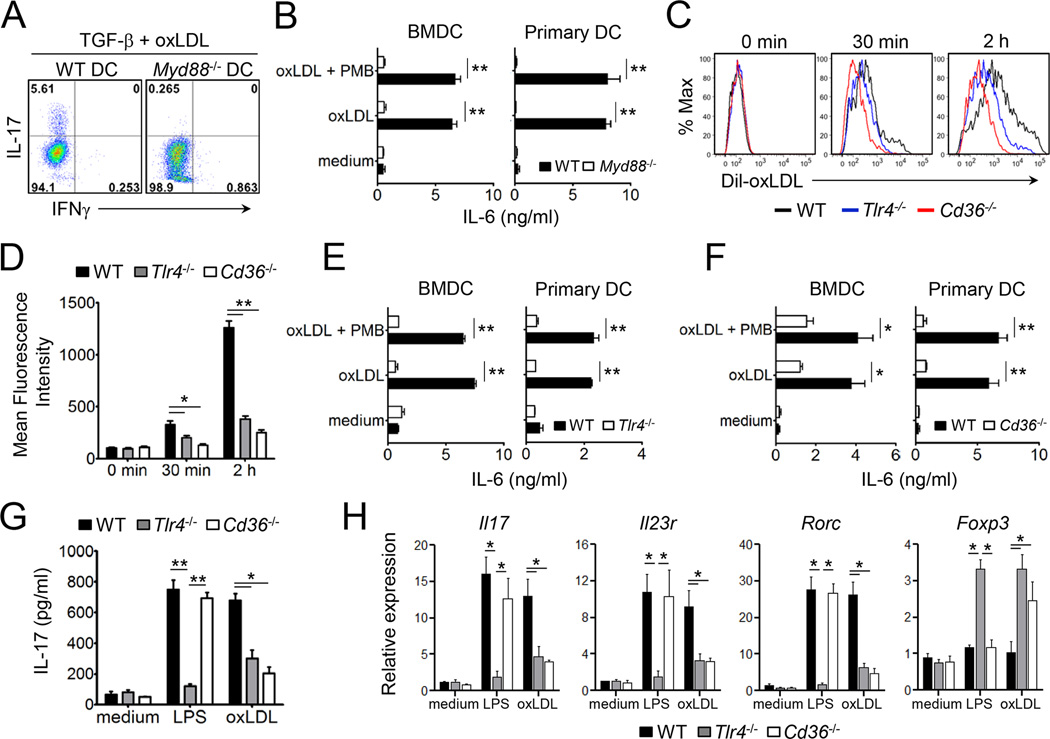
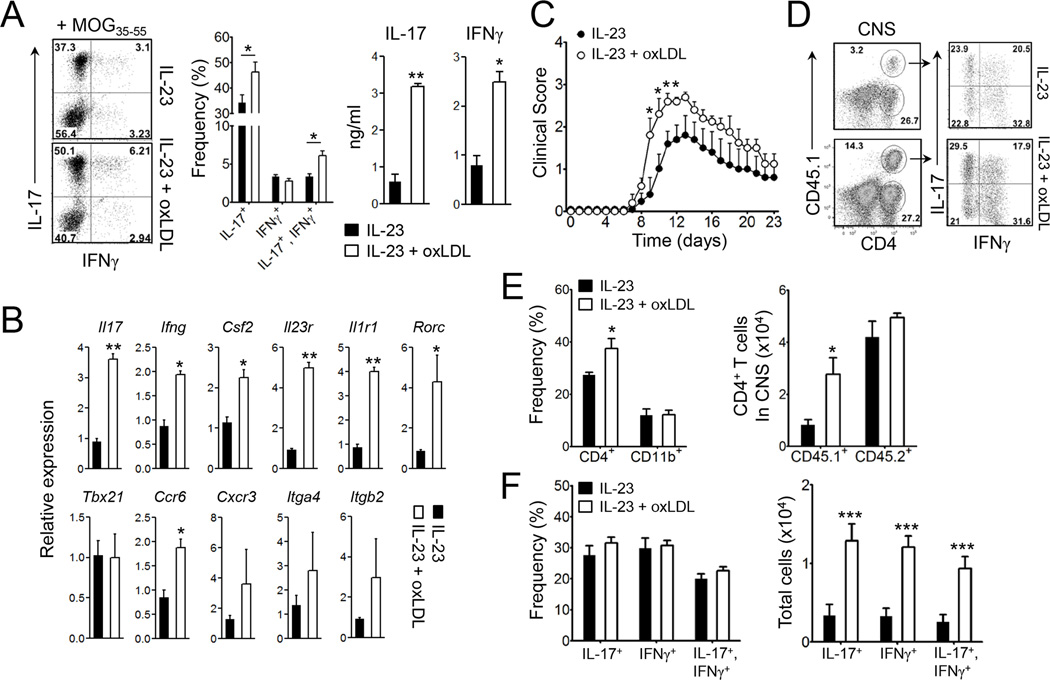
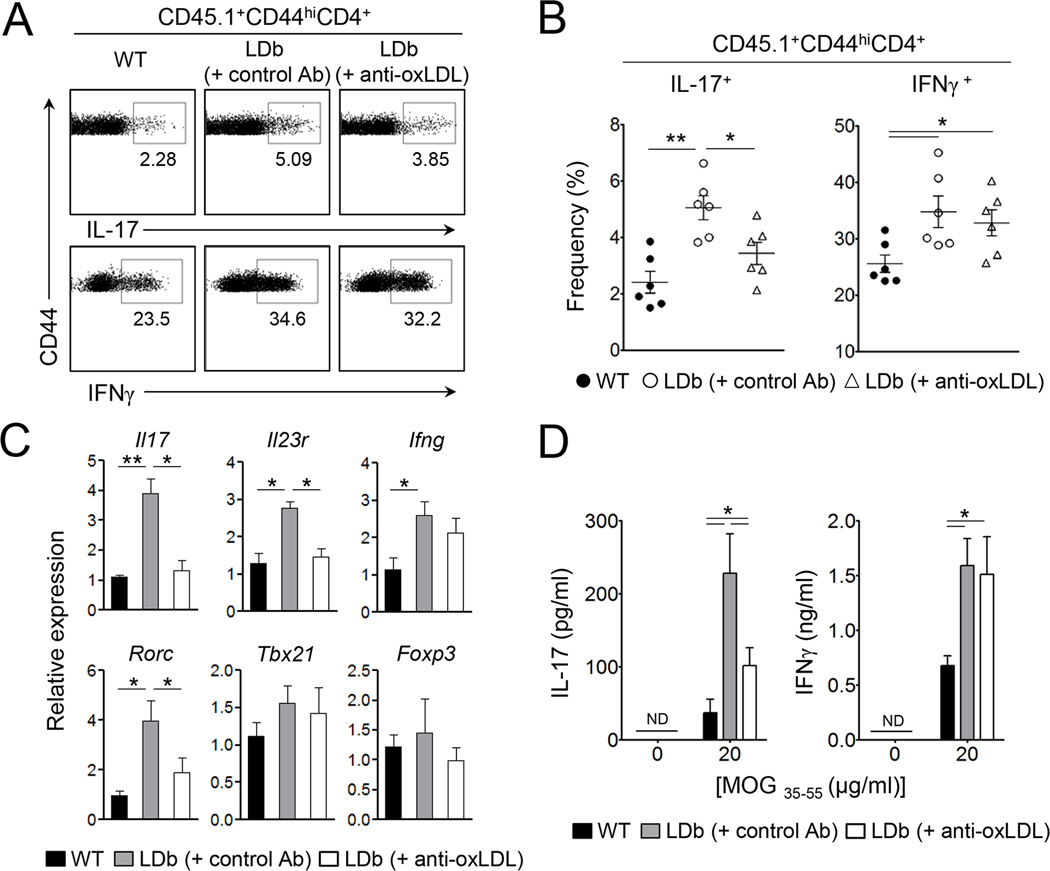
Patients with systemic autoimmune diseases show increased incidence of atherosclerosis. However, the contribution of proatherogenic factors to autoimmunity remains unclear. We found that atherogenic mice (herein referred to as LDb mice) exhibited increased serum interleukin-17, which was associated with increased numbers of T helper 17 (Th17) cells in secondary lymphoid organs. The environment within LDb mice was substantially favorable for Th17 cell polarization of autoreactive T cells during homeostatic proliferation, which was considerably inhibited by antibodies directed against oxidized low-density lipoprotein (oxLDL). Moreover, the uptake of oxLDL induced dendritic-cell-mediated Th17 cell polarization by triggering IL-6 production in a process dependent on TLR4, CD36, and MyD88. Furthermore, self-reactive CD4(+) T cells that expanded in the presence of oxLDL induced more profound experimental autoimmune encephalomyelitis. These findings demonstrate that proatherogenic factors promote the polarization and inflammatory function of autoimmune Th17 cells, which could be critical for the pathogenesis of atherosclerosis and other related autoimmune diseases.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Th17 cells at the crossroads of autoimmunity, inflammation, and atherosclerosis.Immunity. 2014 Jan 16;40(1):10-2. doi: 10.1016/j.immuni.2013.12.006. Immunity. 2014. PMID: 24439264
References
-
- Chawla A, Barak Y, Nagy L, Liao D, Tontonoz P, Evans RM. PPAR-gamma dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat Med. 2001;7:48–52. - PubMed
-
- Danzaki K, Matsui Y, Ikesue M, Ohta D, Ito K, Kanayama M, Kurotaki D, Morimoto J, Iwakura Y, Yagita H, et al. Interleukin-17A deficiency accelerates unstable atherosclerotic plaque formation in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 2012;32:273–280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
