

RNA polymerase II termination involves C-terminal-domain tyrosine dephosphorylation by CPF subunit Glc7
- PMID: 24413056
- PMCID: PMC3917824
- DOI: 10.1038/nsmb.2753
RNA polymerase II termination involves C-terminal-domain tyrosine dephosphorylation by CPF subunit Glc7
Abstract
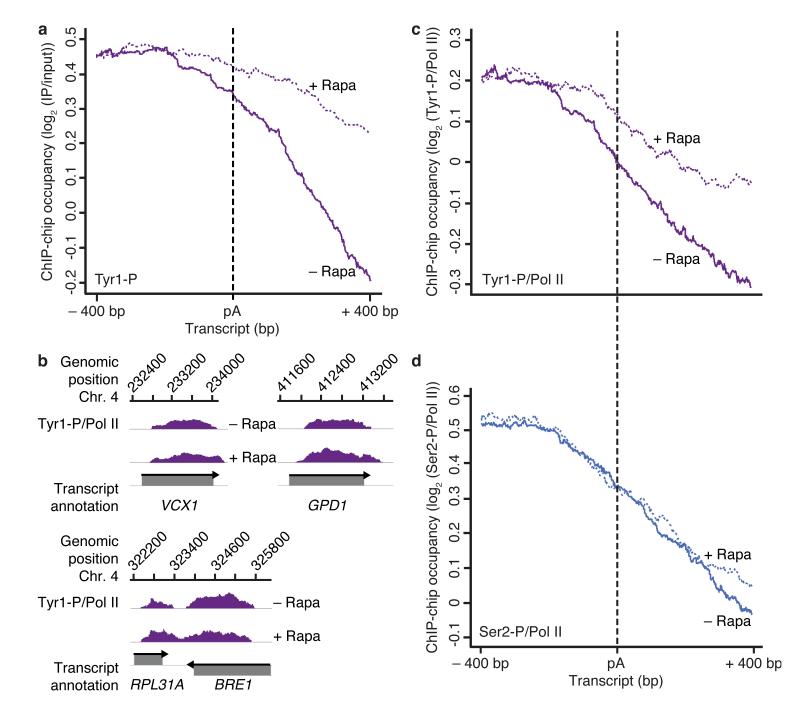
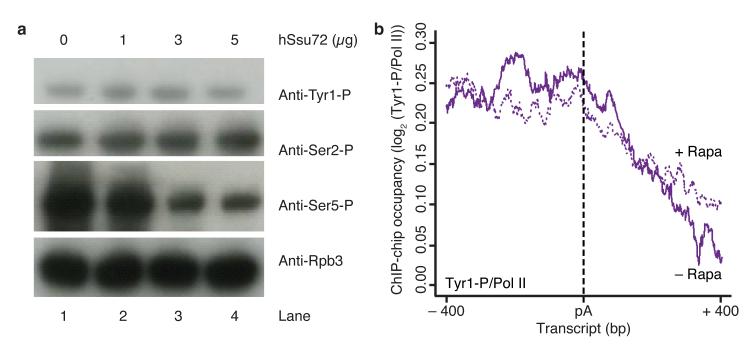
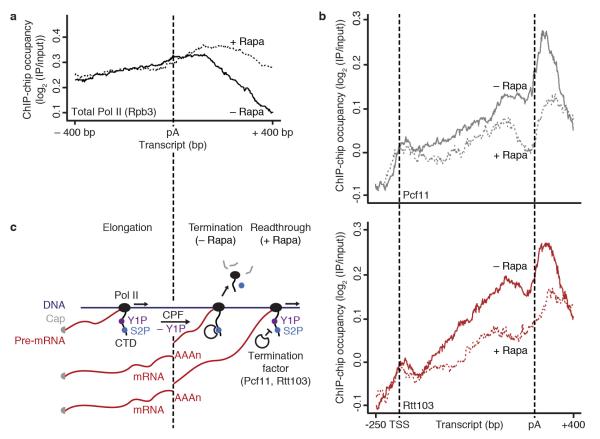
At the 3' ends of protein-coding genes, RNA polymerase (Pol) II is dephosphorylated at tyrosine residues (Tyr1) of its C-terminal domain (CTD). In addition, the associated cleavage-and-polyadenylation factor (CPF) cleaves the transcript and adds a poly(a) tail. Whether these events are coordinated and how they lead to transcription termination remains poorly understood. Here we show that CPF from Saccharomyces cerevisiae is a Pol II-CTD phosphatase and that the CPF subunit Glc7 dephosphorylates Tyr1 in vitro. In vivo, the activity of Glc7 is required for normal Tyr1 dephosphorylation at the polyadenylation site, for recruitment of termination factors Pcf11 and Rtt103 and for normal Pol II termination. These results show that transcription termination involves Tyr1 dephosphorylation of the CTD and indicate that pre-mRNA processing by CPF and transcription termination are coupled via Glc7-dependent Pol II-Tyr1 dephosphorylation.
Figures

References
METHODS REFERENCES
-
- Sydow JF, et al. Structural basis of transcription: mismatch-specific fidelity mechanisms and paused RNA polymerase II with frayed RNA. Mol. Cell. 2009;34:710–721. - PubMed
-
- Chapman RD, et al. Transcribing RNA polymerase II is phosphorylated at CTD residue serine-7. Science. 2007;318:1780–1782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
