

Physical interaction between bacterial heat shock protein (Hsp) 90 and Hsp70 chaperones mediates their cooperative action to refold denatured proteins
- PMID: 24415765
- PMCID: PMC3937677
- DOI: 10.1074/jbc.M113.524801
Physical interaction between bacterial heat shock protein (Hsp) 90 and Hsp70 chaperones mediates their cooperative action to refold denatured proteins
Abstract
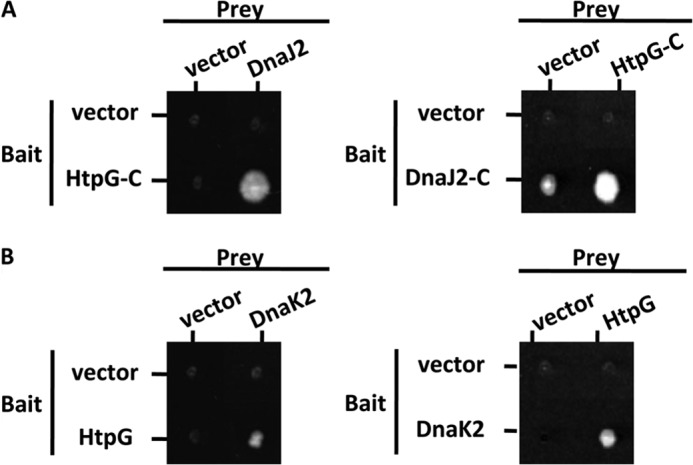
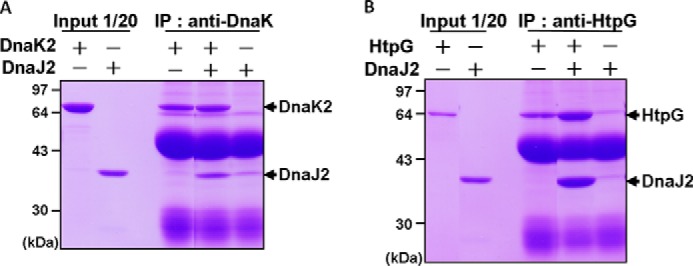
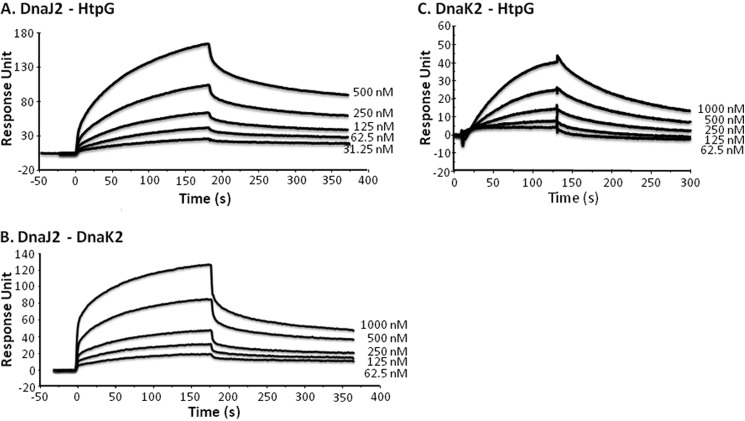
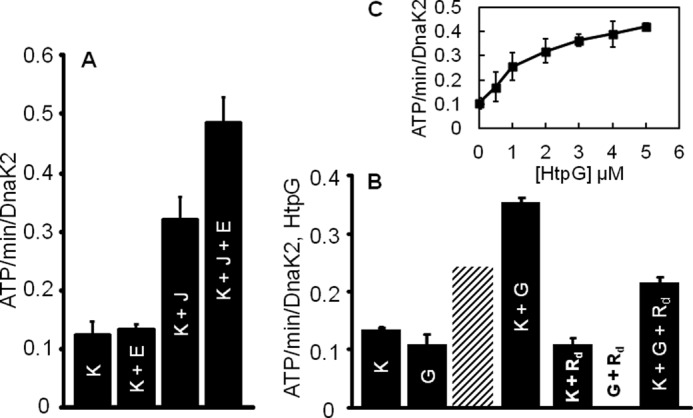
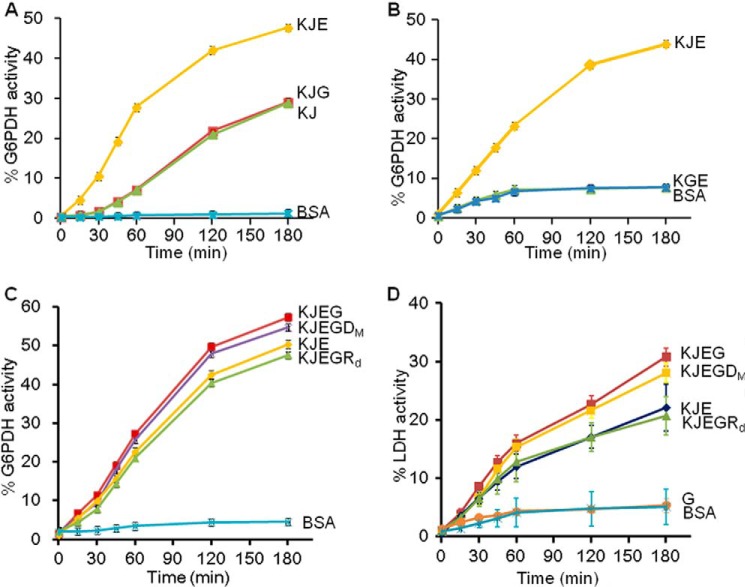
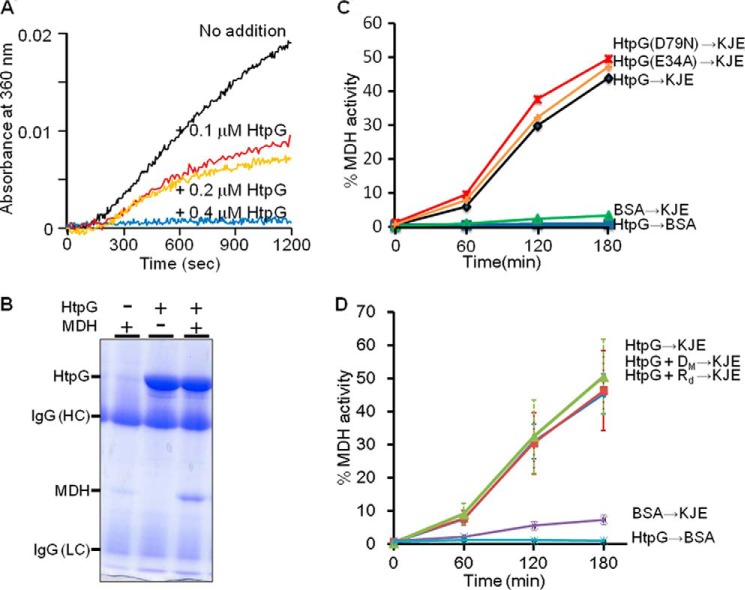
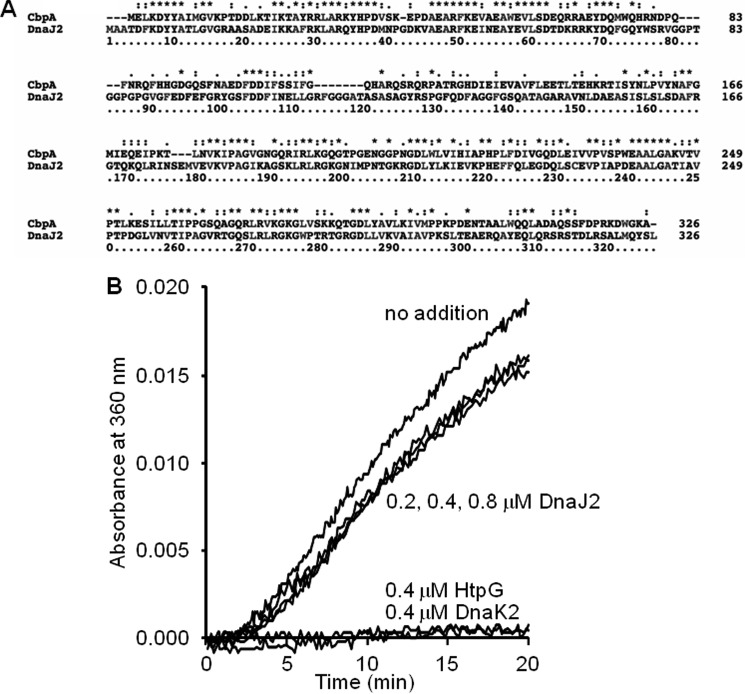
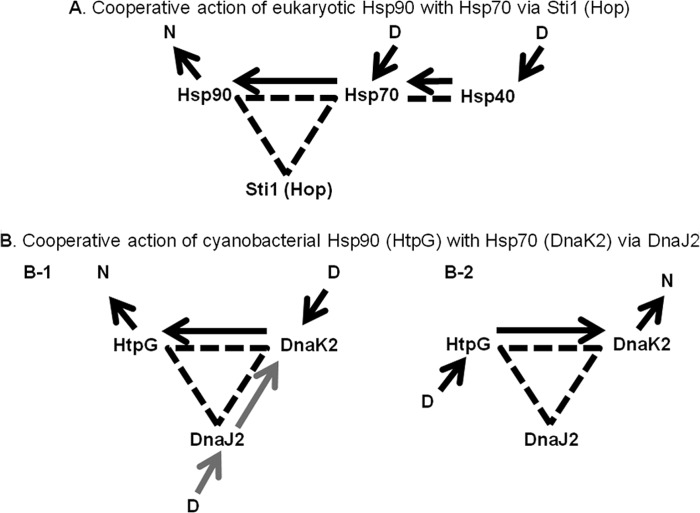
In eukaryotes, heat shock protein 90 (Hsp90) is an essential ATP-dependent molecular chaperone that associates with numerous client proteins. HtpG, a prokaryotic homolog of Hsp90, is essential for thermotolerance in cyanobacteria, and in vitro it suppresses the aggregation of denatured proteins efficiently. Understanding how the non-native client proteins bound to HtpG refold is of central importance to comprehend the essential role of HtpG under stress. Here, we demonstrate by yeast two-hybrid method, immunoprecipitation assays, and surface plasmon resonance techniques that HtpG physically interacts with DnaJ2 and DnaK2. DnaJ2, which belongs to the type II J-protein family, bound DnaK2 or HtpG with submicromolar affinity, and HtpG bound DnaK2 with micromolar affinity. Not only DnaJ2 but also HtpG enhanced the ATP hydrolysis by DnaK2. Although assisted by the DnaK2 chaperone system, HtpG enhanced native refolding of urea-denatured lactate dehydrogenase and heat-denatured glucose-6-phosphate dehydrogenase. HtpG did not substitute for DnaJ2 or GrpE in the DnaK2-assisted refolding of the denatured substrates. The heat-denatured malate dehydrogenase that did not refold by the assistance of the DnaK2 chaperone system alone was trapped by HtpG first and then transferred to DnaK2 where it refolded. Dissociation of substrates from HtpG was either ATP-dependent or -independent depending on the substrate, indicating the presence of two mechanisms of cooperative action between the HtpG and the DnaK2 chaperone system.
Keywords: Cyanobacteria; DnaJ; DnaK; Heat Shock Protein; Hsp70; Hsp90; HtpG; Molecular Chaperone; Protein Aggregation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
