

Notch controls cell adhesion in the Drosophila eye
- PMID: 24415957
- PMCID: PMC3886913
- DOI: 10.1371/journal.pgen.1004087
Notch controls cell adhesion in the Drosophila eye
Abstract
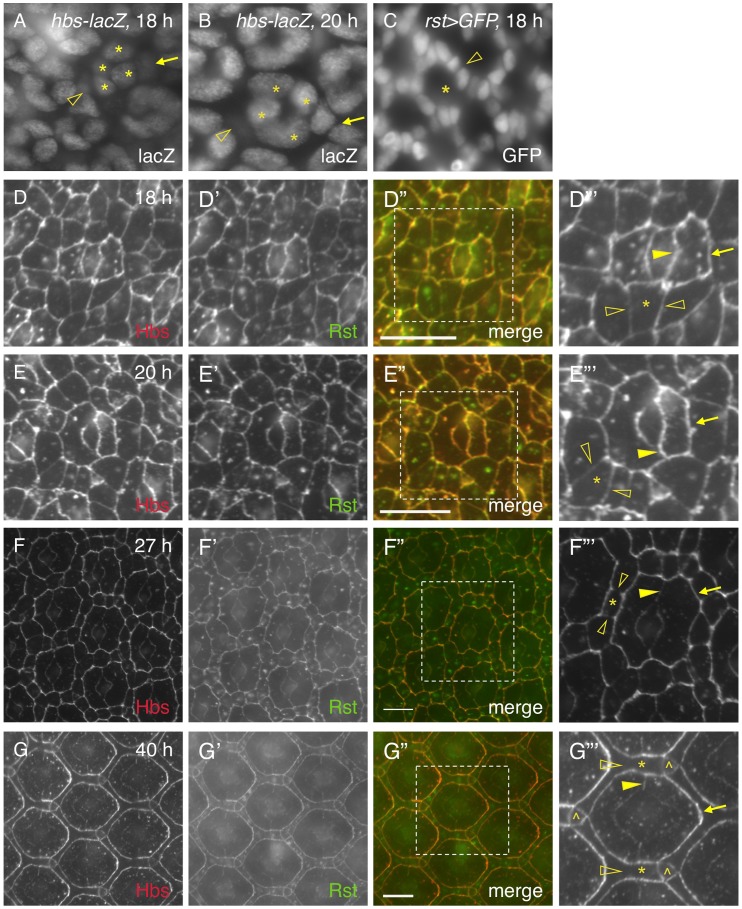
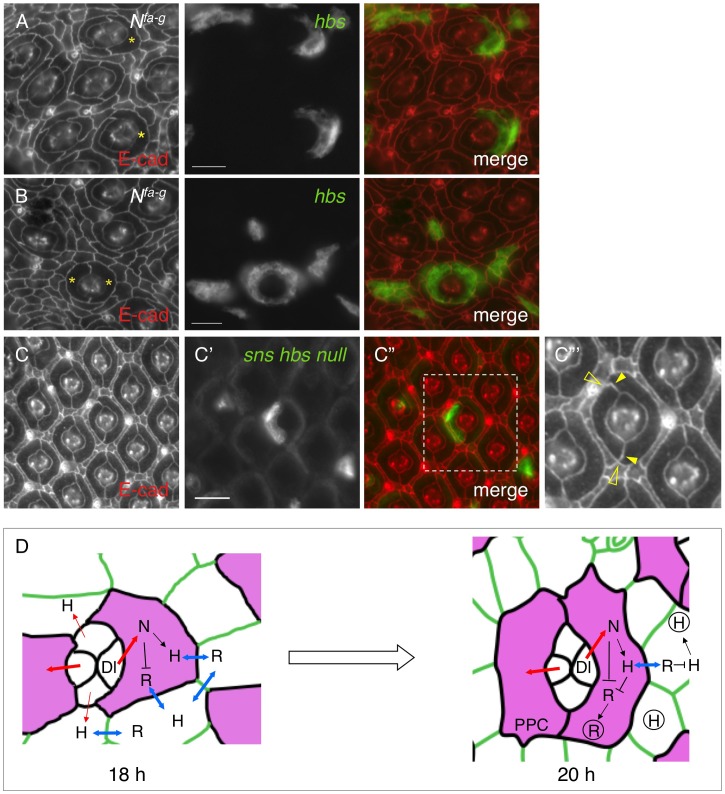
Sporadic evidence suggests Notch is involved in cell adhesion. However, the underlying mechanism is unknown. Here I have investigated an epithelial remodeling process in the Drosophila eye in which two primary pigment cells (PPCs) with a characteristic 'kidney' shape enwrap and eventually isolate a group of cone cells from inter-ommatidial cells (IOCs). This paper shows that in the developing Drosophila eye the ligand Delta was transcribed in cone cells and Notch was activated in the adjacent PPC precursors. In the absence of Notch, emerging PPCs failed to enwrap cone cells, and hibris (hbs) and sns, two genes coding for adhesion molecules of the Nephrin group that mediate preferential adhesion, were not transcribed in PPC precursors. Conversely, activation of Notch in single IOCs led to ectopic expression of hbs and sns. By contrast, in a single IOC that normally transcribes rst, a gene coding for an adhesion molecule of the Neph1 group that binds Hbs and Sns, activation of Notch led to a loss of rst transcription. In addition, in a Notch mutant where two emerging PPCs failed to enwrap cone cells, expression of hbs in PPC precursors restored the ability of these cells to surround cone cells. Further, expression of hbs or rst in a single rst- or hbs-expressing cell, respectively, led to removal of the counterpart from the membrane within the same cell through cis-interaction and forced expression of Rst in all hbs-expressing PPCs strongly disrupted the remodeling process. Finally, a loss of both hbs and sns in single PPC precursors led to constriction of the apical surface that compromised the 'kidney' shape of PPCs. Taken together, these results indicate that cone cells utilize Notch signaling to instruct neighboring PPC precursors to surround them and Notch controls the remodeling process by differentially regulating four adhesion genes.
Conflict of interest statement
The author has declared that no competing interests exist.
Figures





Similar articles
-
A combination of Notch signaling, preferential adhesion and endocytosis induces a slow mode of cell intercalation in the Drosophila retina.Development. 2021 May 15;148(10):dev197301. doi: 10.1242/dev.197301. Epub 2021 May 17. Development. 2021. PMID: 33999996 Free PMC article.
-
Preferential adhesion maintains separation of ommatidia in the Drosophila eye.Dev Biol. 2010 Aug 15;344(2):948-56. doi: 10.1016/j.ydbio.2010.06.013. Epub 2010 Jun 16. Dev Biol. 2010. PMID: 20599904 Free PMC article.
-
The immunoglobulin superfamily member Hbs functions redundantly with Sns in interactions between founder and fusion-competent myoblasts.Development. 2009 Apr;136(7):1159-68. doi: 10.1242/dev.026302. Development. 2009. PMID: 19270174 Free PMC article.
-
Pattern formation in the Drosophila eye.Curr Opin Genet Dev. 2007 Aug;17(4):309-13. doi: 10.1016/j.gde.2007.05.001. Epub 2007 Jul 6. Curr Opin Genet Dev. 2007. PMID: 17618111 Free PMC article. Review.
-
Robust selection of sensory organ precursors by the Notch-Delta pathway.Curr Opin Cell Biol. 2011 Dec;23(6):663-7. doi: 10.1016/j.ceb.2011.09.005. Epub 2011 Sep 29. Curr Opin Cell Biol. 2011. PMID: 21963301 Review.
Cited by
-
Notch and Delta Control the Switch and Formation of Camouflage Patterns in Caterpillars.iScience. 2020 Jul 24;23(7):101315. doi: 10.1016/j.isci.2020.101315. Epub 2020 Jun 25. iScience. 2020. PMID: 32650115 Free PMC article.
-
Notch Target Gene E(spl)mδ Is a Mediator of Methylmercury-Induced Myotoxicity in Drosophila.Front Genet. 2018 Jan 15;8:233. doi: 10.3389/fgene.2017.00233. eCollection 2017. Front Genet. 2018. PMID: 29379520 Free PMC article.
-
Hexagonal patterning of the Drosophila eye.Dev Biol. 2021 Oct;478:173-182. doi: 10.1016/j.ydbio.2021.07.004. Epub 2021 Jul 8. Dev Biol. 2021. PMID: 34245727 Free PMC article. Review.
-
A combination of Notch signaling, preferential adhesion and endocytosis induces a slow mode of cell intercalation in the Drosophila retina.Development. 2021 May 15;148(10):dev197301. doi: 10.1242/dev.197301. Epub 2021 May 17. Development. 2021. PMID: 33999996 Free PMC article.
-
Neuronal immunoglobulin superfamily cell adhesion molecules in epithelial morphogenesis: insights from Drosophila.Philos Trans R Soc Lond B Biol Sci. 2020 Oct 12;375(1809):20190553. doi: 10.1098/rstb.2019.0553. Epub 2020 Aug 24. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32829687 Free PMC article. Review.
References
-
- Barolo S, Posakony JW (2002) Three habits of highly effective signaling pathways: principles of transcriptional control by developmental cell signaling. Genes Dev 16: 1167–1181. - PubMed
-
- Dahmann C, Basler K (1999) Compartment boundaries: at the edge of development. Trends Genet 15: 320–326. - PubMed
-
- Irvine KD, Vogt TF (1997) Dorsal-ventral signaling in limb development. Curr Opin Cell Biol 9: 867–876. - PubMed
-
- Cagan RL, Ready DF (1989) Notch is required for successive cell decisions in the developing Drosophila retina. Genes Dev 3: 1099–1112. - PubMed
-
- Hoppe PE, Greenspan RJ (1986) Local function of the Notch gene for embryonic ectodermal pathway choice in Drosophila. Cell 46: 773–783. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
