

Transcriptional and epigenetic regulation of Hebbian and non-Hebbian plasticity
- PMID: 24418102
- PMCID: PMC3984596
- DOI: 10.1016/j.neuropharm.2014.01.001
Transcriptional and epigenetic regulation of Hebbian and non-Hebbian plasticity
Abstract
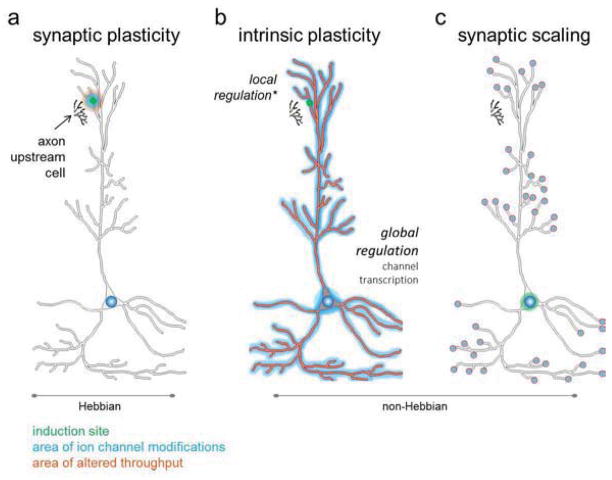
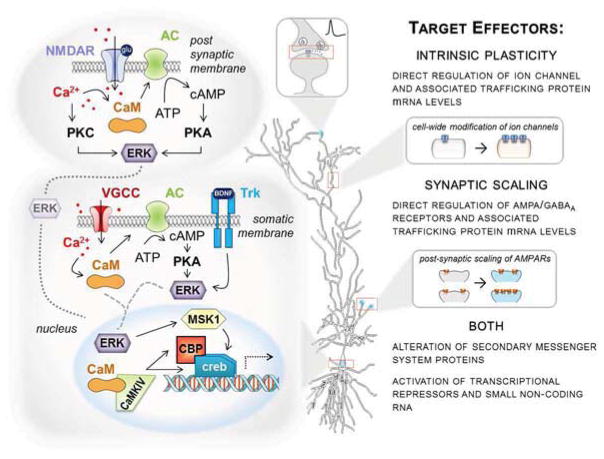
The epigenome is uniquely positioned as a point of convergence, integrating multiple intracellular signaling cascades into a cohesive gene expression profile necessary for long-term behavioral change. The last decade of neuroepigenetic research has primarily focused on learning-induced changes in DNA methylation and chromatin modifications. Numerous studies have independently demonstrated the importance of epigenetic modifications in memory formation and retention as well as Hebbian plasticity. However, how these mechanisms operate in the context of other forms of plasticity is largely unknown. In this review, we examine evidence for epigenetic regulation of Hebbian plasticity. We then discuss how non-Hebbian forms of plasticity, such as intrinsic plasticity and synaptic scaling, may also be involved in producing the cellular adaptations necessary for learning-related behavioral change. Furthermore, we consider the likely roles for transcriptional and epigenetic mechanisms in the regulation of these plasticities. In doing so, we aim to expand upon the idea that epigenetic mechanisms are critical regulators of both Hebbian and non-Hebbian forms of plasticity that ultimately drive learning and memory.
Keywords: DNA methylation; Epigenetics; Hebbian; Histone modifications; Homeostatic; Intrinsic; Metaplasticity; Non-Hebbian; Synaptic; Synaptic scaling.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Alarcon JMJ, Malleret GG, Touzani KK, Vronskaya SS, Ishii SS, Kandel ERE, Barco AA. Chromatin Acetylation, Memory, and LTP Are Impaired in CBP^+^/^- Mice - A Model for the Cognitive Deficit in Rubinstein-Taybi Syndrome and Its Amelioration. Neuron. 2004;42:13–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
