

Coordination between zinc and phosphate homeostasis involves the transcription factor PHR1, the phosphate exporter PHO1, and its homologue PHO1;H3 in Arabidopsis
- PMID: 24420568
- PMCID: PMC3924728
- DOI: 10.1093/jxb/ert444
Coordination between zinc and phosphate homeostasis involves the transcription factor PHR1, the phosphate exporter PHO1, and its homologue PHO1;H3 in Arabidopsis
Abstract
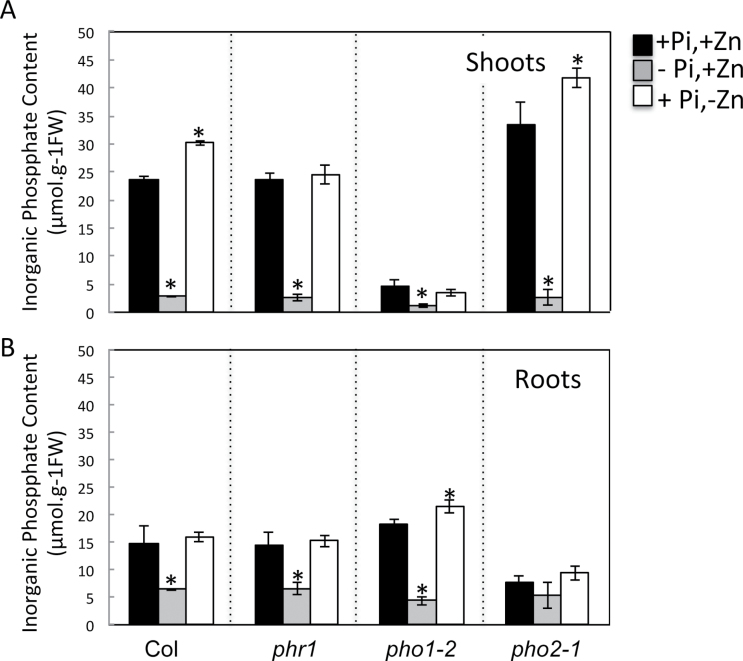
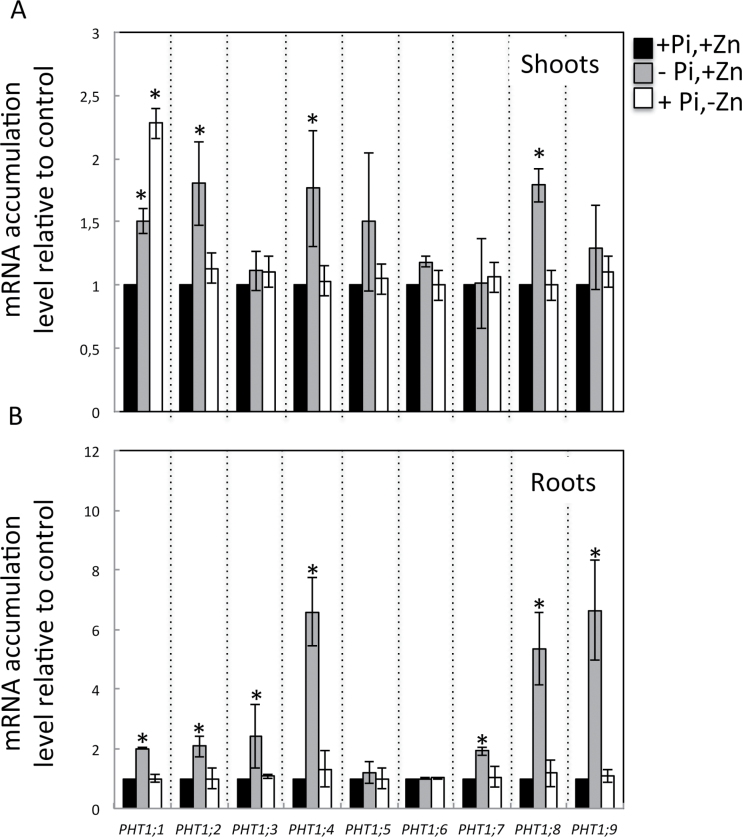
Interactions between zinc (Zn) and phosphate (Pi) nutrition in plants have long been recognized, but little information is available on their molecular bases and biological significance. This work aimed at examining the effects of Zn deficiency on Pi accumulation in Arabidopsis thaliana and uncovering genes involved in the Zn-Pi synergy. Wild-type plants as well as mutants affected in Pi signalling and transport genes, namely the transcription factor PHR1, the E2-conjugase PHO2, and the Pi exporter PHO1, were examined. Zn deficiency caused an increase in shoot Pi content in the wild type as well as in the pho2 mutant, but not in the phr1 or pho1 mutants. This indicated that PHR1 and PHO1 participate in the coregulation of Zn and Pi homeostasis. Zn deprivation had a very limited effect on transcript levels of Pi-starvation-responsive genes such as AT4, IPS1, and microRNA399, or on of members of the high-affinity Pi transporter family PHT1. Interestingly, one of the PHO1 homologues, PHO1;H3, was upregulated in response to Zn deficiency. The expression pattern of PHO1 and PHO1;H3 were similar, both being expressed in cells of the root vascular cylinder and both localized to the Golgi when expressed transiently in tobacco cells. When grown in Zn-free medium, pho1;h3 mutant plants displayed higher Pi contents in the shoots than wild-type plants. This was, however, not observed in a pho1 pho1;h3 double mutant, suggesting that PHO1;H3 restricts root-to-shoot Pi transfer requiring PHO1 function for Pi homeostasis in response to Zn deficiency.
Keywords: Homeostasis; interaction; phosphate; signalling; transport; zinc..
Figures







References
-
- Alonso JM, Stepanova AN, Leisse TJ, et al. , 2003. Genome-wide insertional mutagenesis of Arabidopsis thaliana . Science 301, 653–657 - PubMed
-
- Ames BN. 1966. Assay of inorganic phosphate, total phosphate and phosphatases. Methods in Enzymology 8, 115–118
-
- Arpat AB, Magliano P, Wege S, Rouached H, Stefanovic A, Poirier Y. 2012. Functional expression of PHO1 to the Golgi and trans-Golgi network and its role in export of inorganic phosphate. The Plant Journal 71, 479–491 - PubMed
-
- Batistic O, Waadt R, Steinhorst L, Held K, Kudla J. 2010. CBL-mediated targeting of CIPKs facilitates the decoding of calcium signals emanating from distinct cellular stores. The Plant Journal 61, 211–222 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
