

Functional regeneration beyond the glial scar
- PMID: 24424280
- PMCID: PMC3951813
- DOI: 10.1016/j.expneurol.2013.12.024
Functional regeneration beyond the glial scar
Abstract
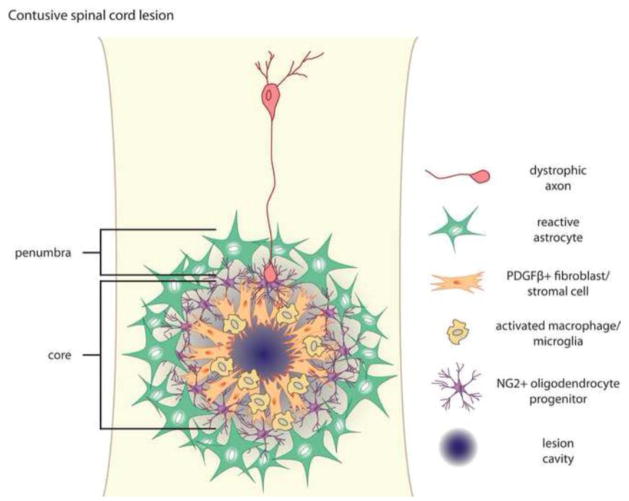
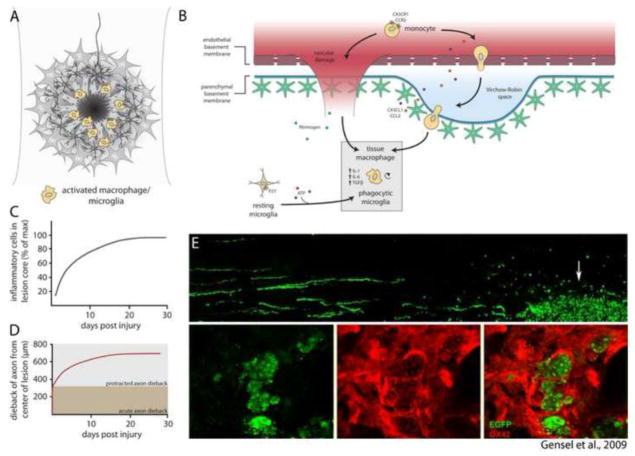
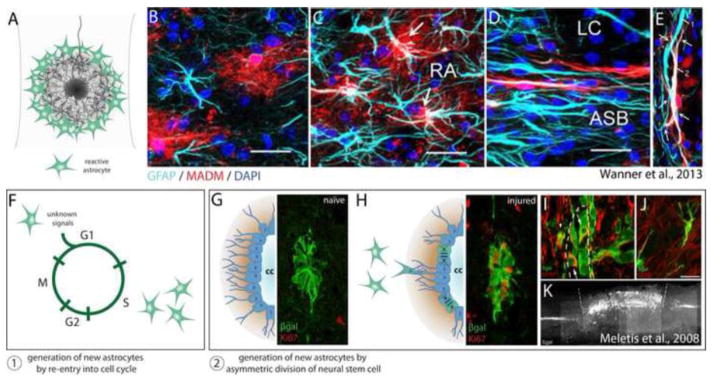
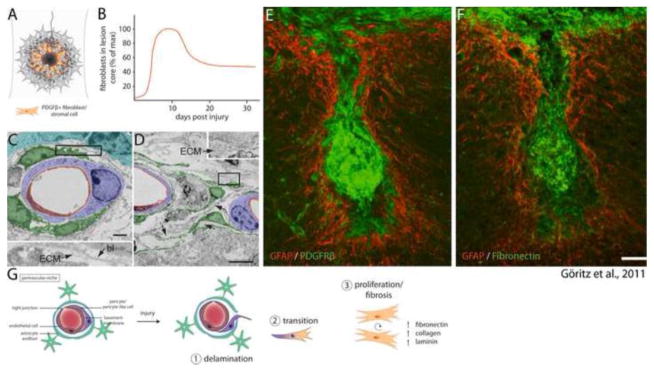
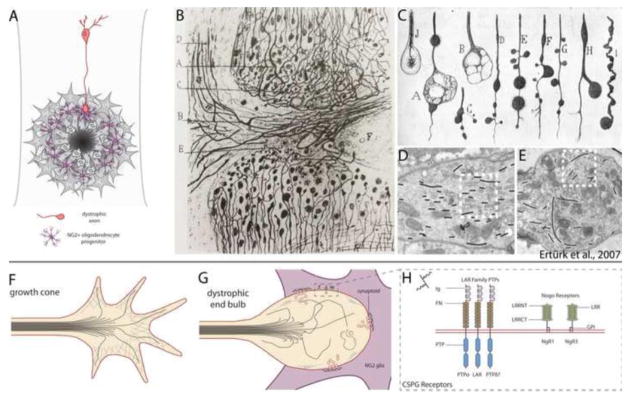
Astrocytes react to CNS injury by building a dense wall of filamentous processes around the lesion. Stromal cells quickly take up residence in the lesion core and synthesize connective tissue elements that contribute to fibrosis. Oligodendrocyte precursor cells proliferate within the lesion and entrap dystrophic axon tips. Here we review evidence that this aggregate scar acts as the major barrier to regeneration of axons after injury. We also consider several exciting new interventions that allow axons to regenerate beyond the glial scar, and discuss the implications of this work for the future of regeneration biology.
Keywords: Axon growth cone; Chondroitin sulfate proteoglycans; Glial scar; Hypertrophy; Regeneration; Spinal cord injury.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Ajami B, Bennett JL, Krieger C, McNagny KM, Rossi FM. Infiltrating monocytes trigger EAE progression, but do not contribute to the resident microglia pool. Nat Neurosci. 2011;14:1142–1149. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
