

Serines 440 and 467 in the Werner syndrome protein are phosphorylated by DNA-PK and affects its dynamics in response to DNA double strand breaks
- PMID: 24429382
- PMCID: PMC3927811
- DOI: 10.18632/aging.100629
Serines 440 and 467 in the Werner syndrome protein are phosphorylated by DNA-PK and affects its dynamics in response to DNA double strand breaks
Abstract
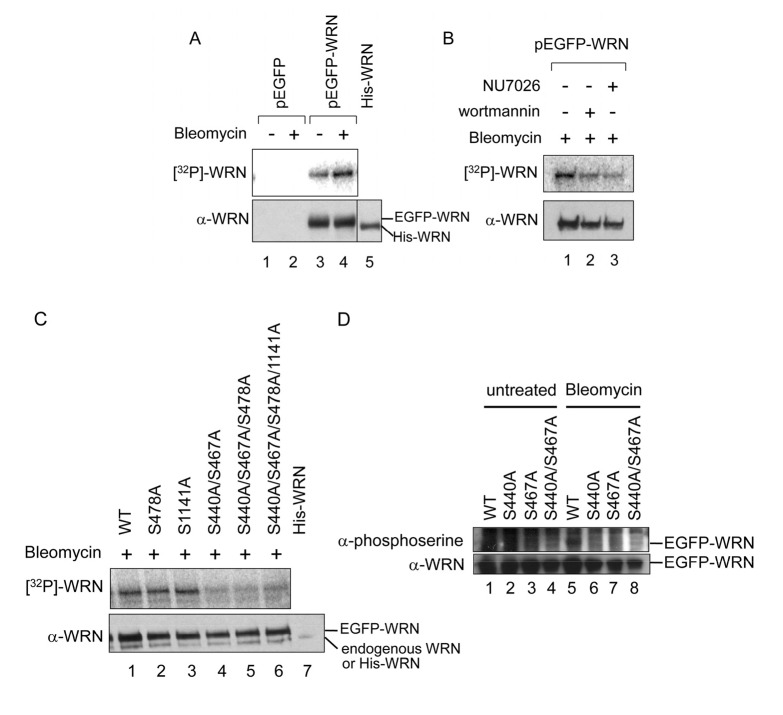
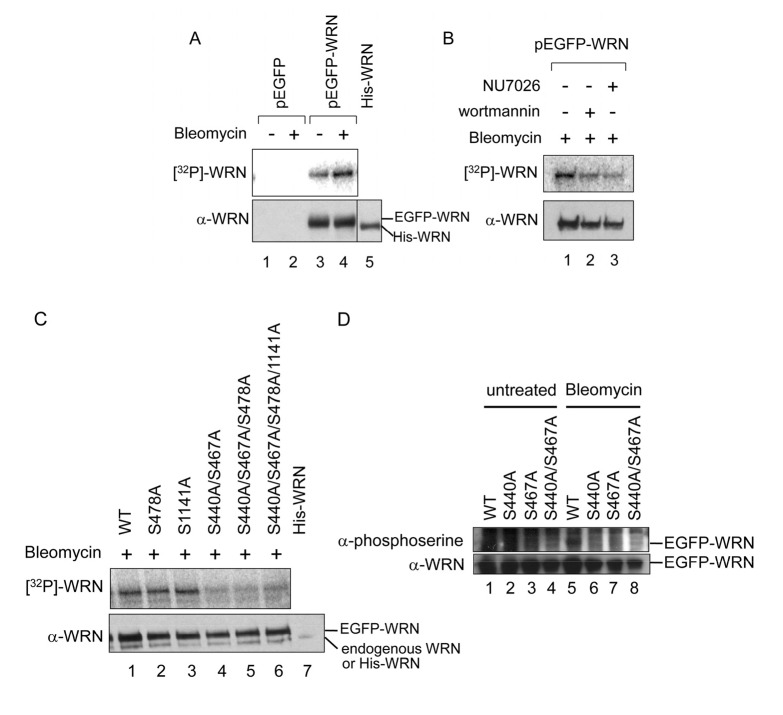
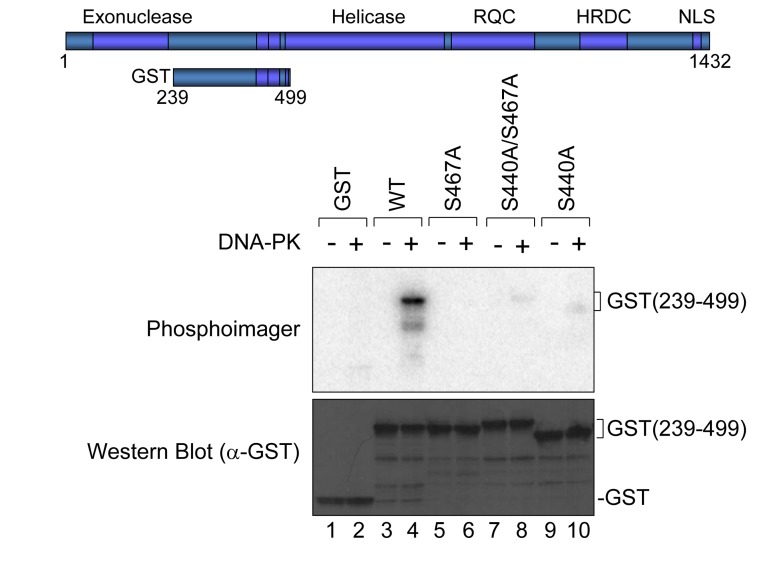
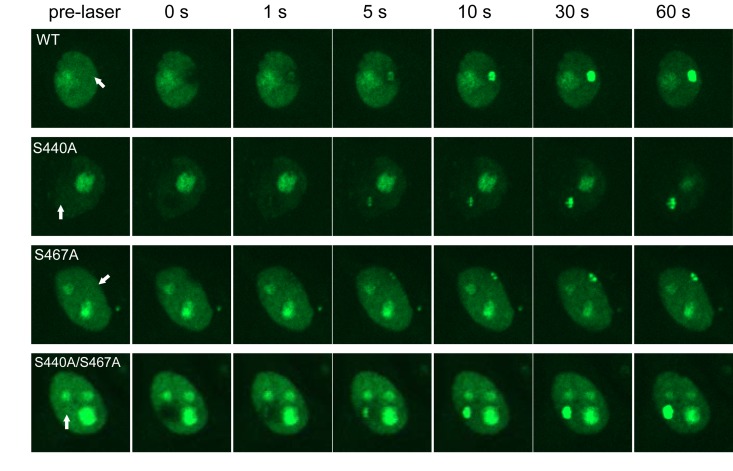
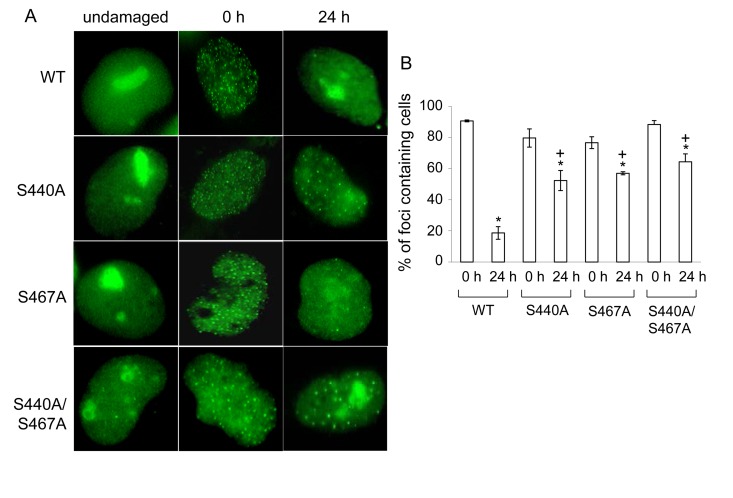
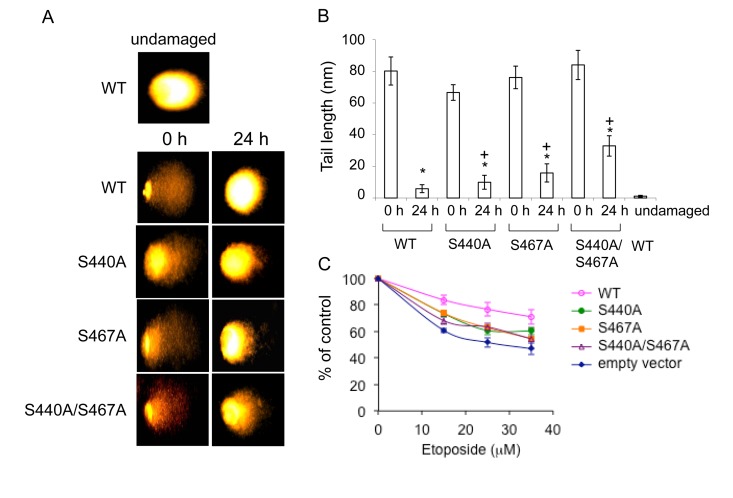
WRN protein, defective in Werner syndrome (WS), a human segmental progeria, is a target of serine/threonine kinases involved in sensing DNA damage. DNA-PK phosphorylates WRN in response to DNA double strand breaks (DSBs). However, the main phosphorylation sites and functional importance of the phosphorylation of WRN has remained unclear. Here, we identify Ser-440 and -467 in WRN as major phosphorylation sites mediated by DNA-PK.In vitro, DNA-PK fails to phosphorylate a GST-WRN fragment with S440A and/or S467A substitution. In addition, full length WRN with the mutation expressed in 293T cells was not phosphorylated in response to DSBs produced by bleomycin. Accumulation of the mutant WRN at the site of laser-induced DSBs occurred with the same kinetics as wild type WRN in live HeLa cells. While the wild type WRN relocalized to the nucleoli after 24 hours recovery from etoposide-induced DSBs, the mutant WRN remained mostly in the nucleoplasm. Consistent with this, WS cells expressing the mutants exhibited less DNA repair efficiency and more sensitivity to etoposide, compared to those expressing wild type. Our findings indicate that phosphorylation of Ser-440 and -467 in WRN are important for relocalization of WRN to nucleoli, and that it is required for efficient DSB repair.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Goto M. Werner's syndrome: from clinics to genetics. Clin Exp Rheumatol. 2000;18:760–766. - PubMed
-
- Martin GM, Sprague CA, Epstein CJ. Replicative life-span of cultivated human cells. Effects of donor's age, tissue, and genotype. Lab Invest. 1970;23:86–92. - PubMed
-
- Salk D, Au K, Hoehn H, Martin GM. Cytogenetics of Werner's syndrome cultured skin fibroblasts: variegated translocation mosaicism. Cytogenet Cell Genet. 1981;30:92–107. - PubMed
-
- Yannone SM, Roy S, Chan DW, Murphy MB, Huang S, Campisi J, et al. Werner syndrome protein is regulated and phosphorylated by DNA-dependent protein kinase. J Biol Chem. 2001;276:38242–38248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
