

Therapeutic inflammatory monocyte modulation using immune-modifying microparticles
- PMID: 24431111
- PMCID: PMC3973033
- DOI: 10.1126/scitranslmed.3007563
Therapeutic inflammatory monocyte modulation using immune-modifying microparticles
Abstract
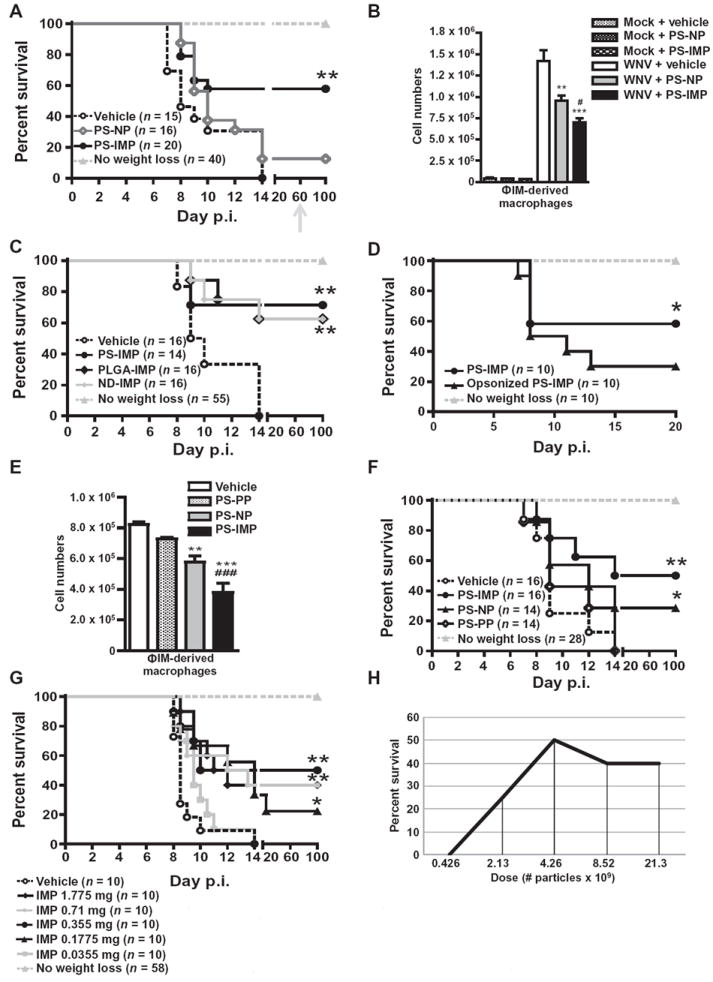
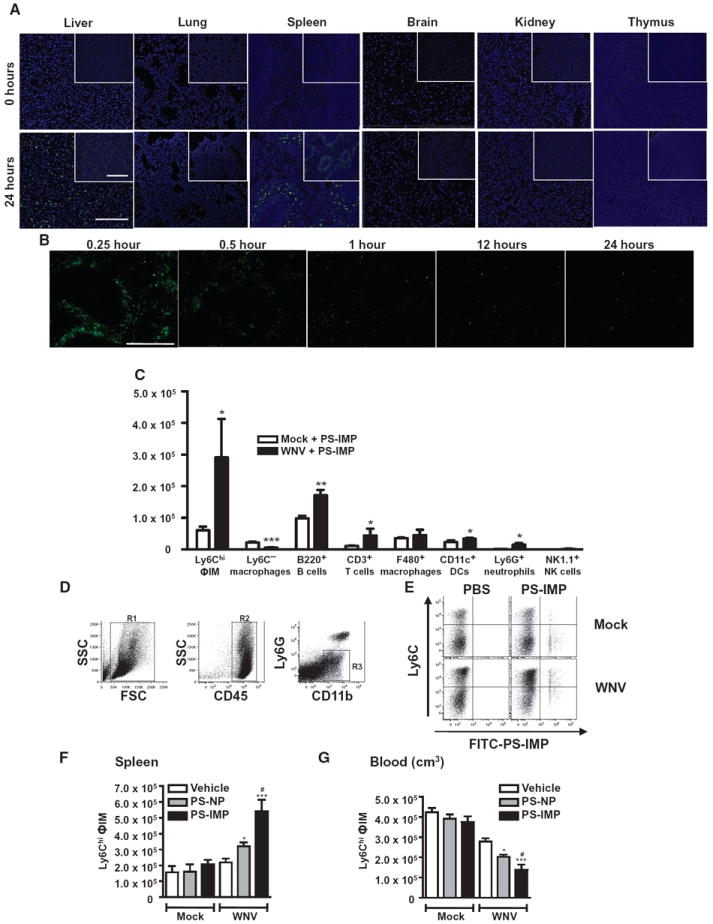
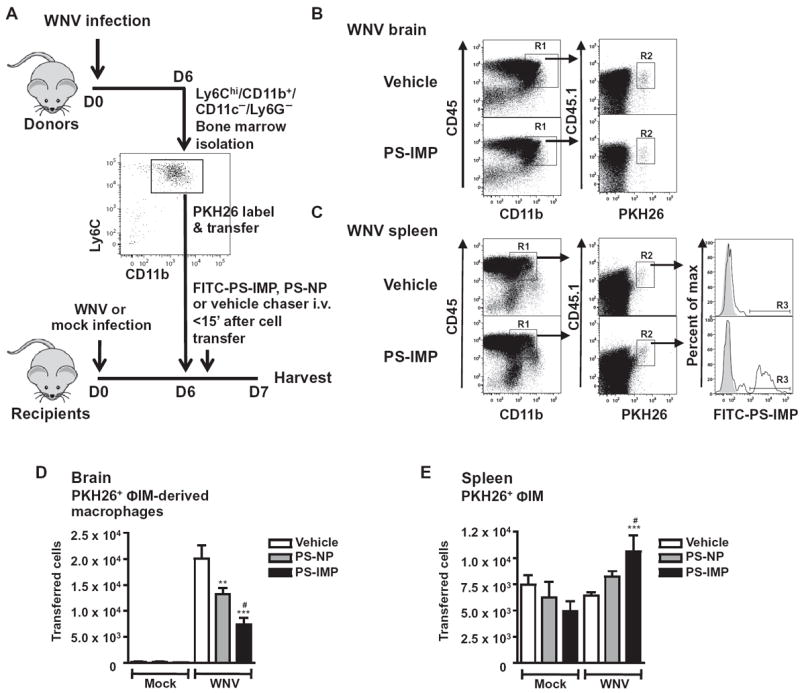
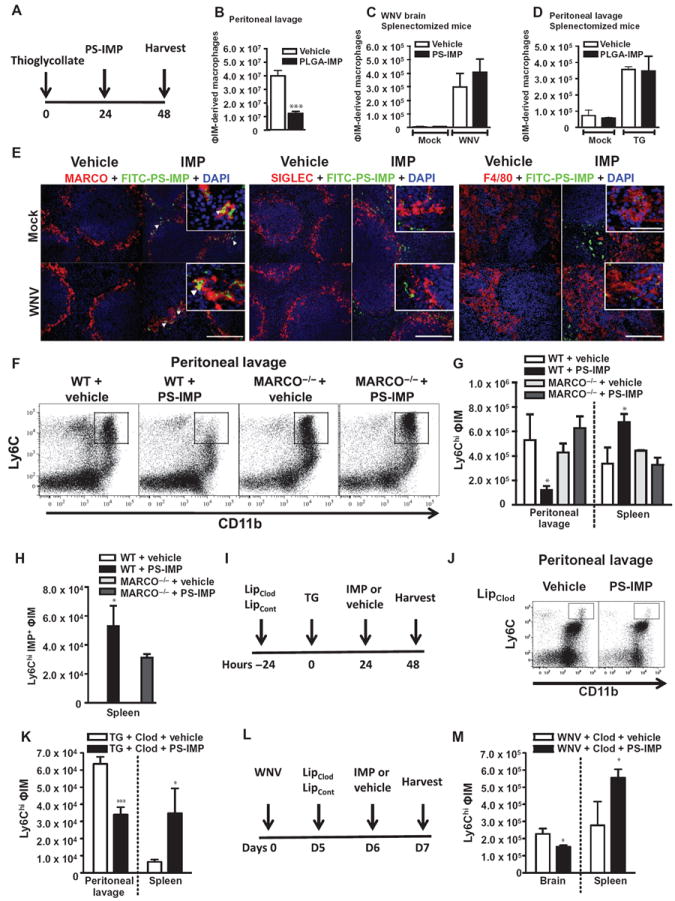
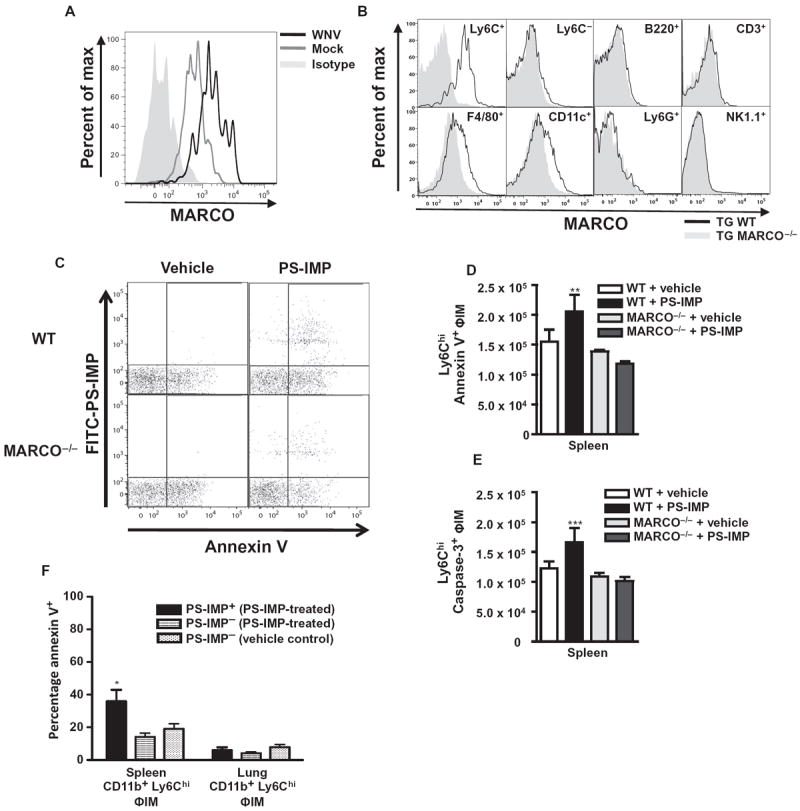
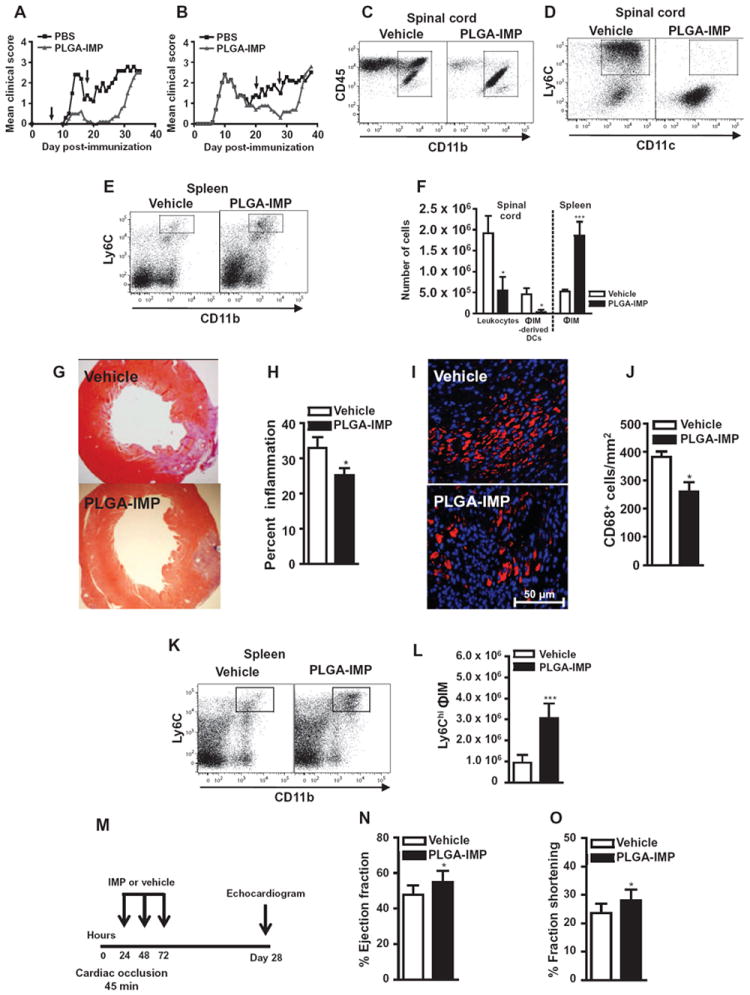
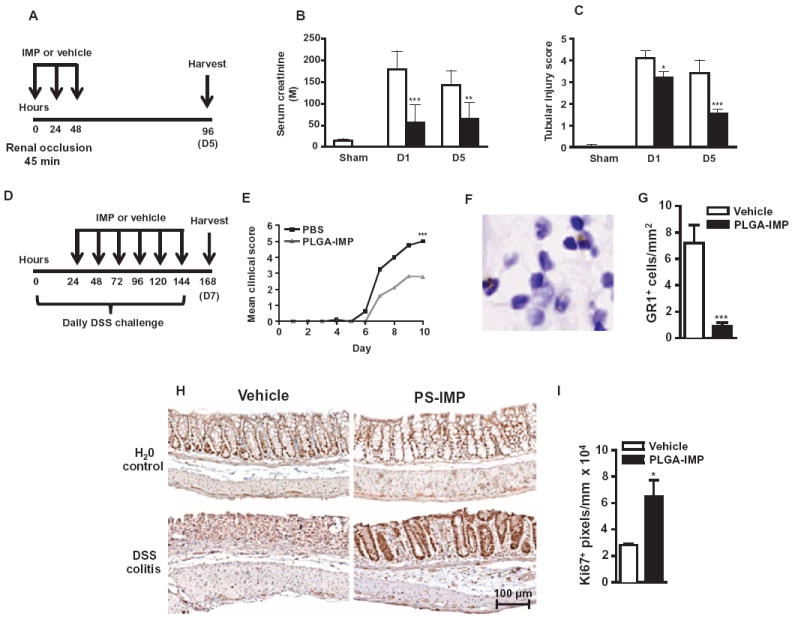
Inflammatory monocyte-derived effector cells play an important role in the pathogenesis of numerous inflammatory diseases. However, no treatment option exists that is capable of modulating these cells specifically. We show that infused negatively charged, immune-modifying microparticles (IMPs), derived from polystyrene, microdiamonds, or biodegradable poly(lactic-co-glycolic) acid, were taken up by inflammatory monocytes, in an opsonin-independent fashion, via the macrophage receptor with collagenous structure (MARCO). Subsequently, these monocytes no longer trafficked to sites of inflammation; rather, IMP infusion caused their sequestration in the spleen through apoptotic cell clearance mechanisms and, ultimately, caspase-3-mediated apoptosis. Administration of IMPs in mouse models of myocardial infarction, experimental autoimmune encephalomyelitis, dextran sodium sulfate-induced colitis, thioglycollate-induced peritonitis, and lethal flavivirus encephalitis markedly reduced monocyte accumulation at inflammatory foci, reduced disease symptoms, and promoted tissue repair. Together, these data highlight the intricate interplay between scavenger receptors, the spleen, and inflammatory monocyte function and support the translation of IMPs for therapeutic use in diseases caused or potentiated by inflammatory monocytes.
Conflict of interest statement
Figures
Comment in
-
Inflammatory disorders: monocytes derailed by microparticles.Nat Rev Drug Discov. 2014 Mar;13(3):175. doi: 10.1038/nrd4263. Epub 2014 Feb 14. Nat Rev Drug Discov. 2014. PMID: 24525780 No abstract available.
References
-
- Bain CC, Scott CL, Uronen-Hansson H, Gudjonsson S, Jansson O, Grip O, Guilliams M, Malissen B, Agace WW, Mowat AM. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 2013;6:498–510. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
