

Photoinactivation of glutamate receptors by genetically encoded unnatural amino acids
- PMID: 24431456
- PMCID: PMC6608342
- DOI: 10.1523/JNEUROSCI.3725-13.2014
Photoinactivation of glutamate receptors by genetically encoded unnatural amino acids
Abstract
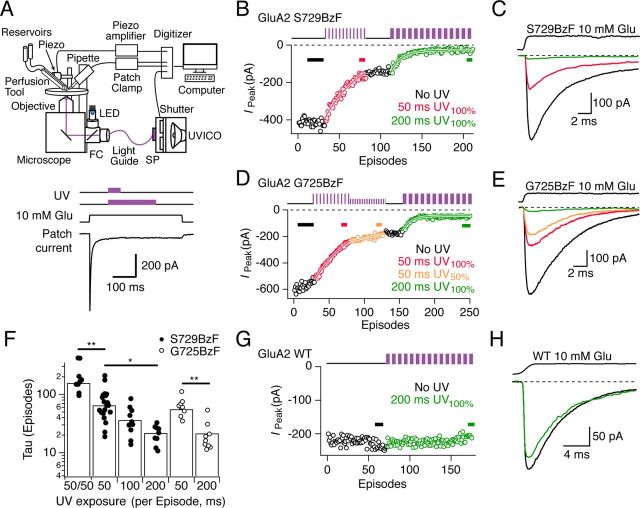
Ionotropic glutamate receptors (iGluRs) are ubiquitous in the mammalian brain, and the AMPA-subtype is essential for fast, glutamate-activated postsynaptic currents. We incorporated photoactive crosslinkers into AMPA receptors using genetically encoded unnatural amino acid mutagenesis in a mammalian cell line. Receptors rescued by incorporation of unnatural amino acids, including p-benzoyl-l-phenylalanine (BzF, also known as Bpa), had largely similar properties to wild-type channels and were expressed at similar levels. BzF incorporation at subunit interfaces afforded photocrosslinking of subunits, as assessed by biochemical experiments. In electrophysiological recordings, BzF incorporation allowed selective and potent UV-driven photoinactivation of both homomeric (GluA2) and heteromeric (GluA2:GluA1) AMPA receptors. State dependence of trapping at two sites in the lower lobe of the ligand binding domain is consistent with deformation of these domains as well as intersubunit rearrangements during AMPA receptor desensitization.
Keywords: UV light; benzophenone; crosslinking; mammalian cells; orthogonal tRNA; synthetase.
Figures







References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources