

Fibre type composition in the lumbar perivertebral muscles of primates: implications for the evolution of orthogrady in hominoids
- PMID: 24433382
- PMCID: PMC3969056
- DOI: 10.1111/joa.12130
Fibre type composition in the lumbar perivertebral muscles of primates: implications for the evolution of orthogrady in hominoids
Abstract
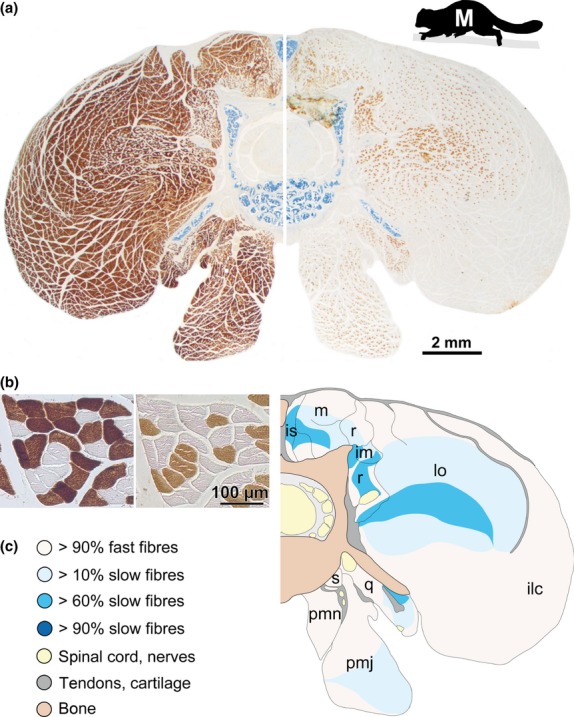
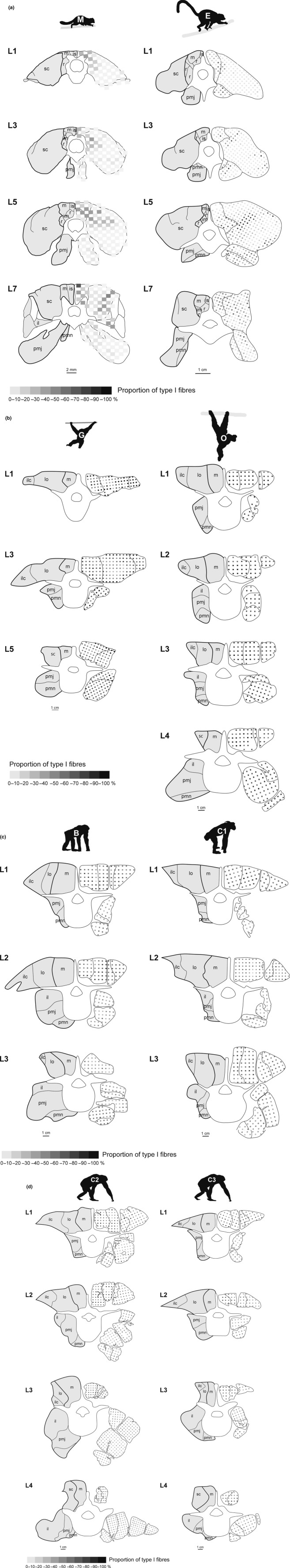
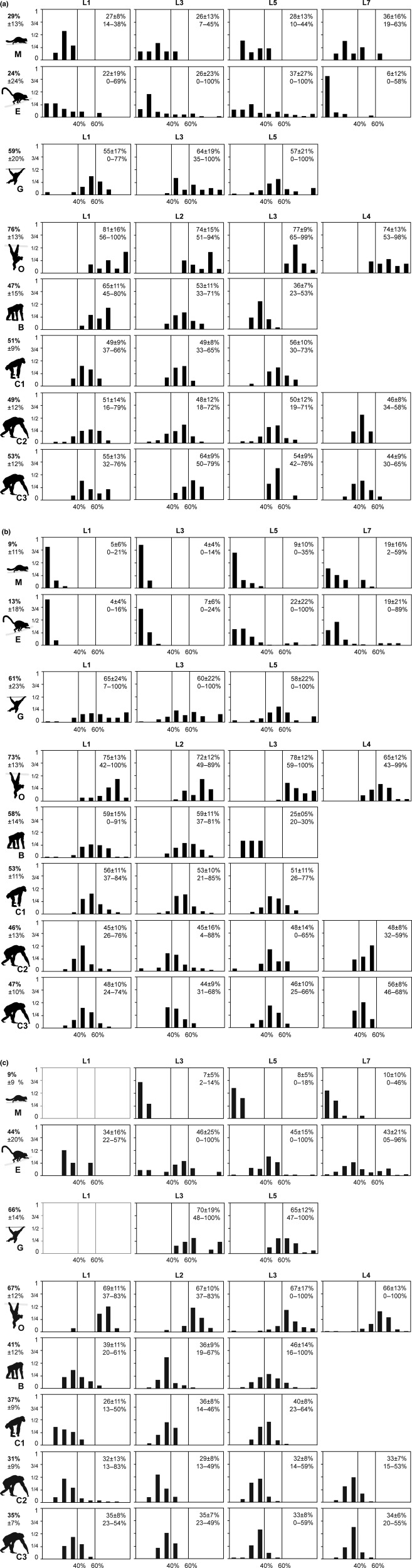
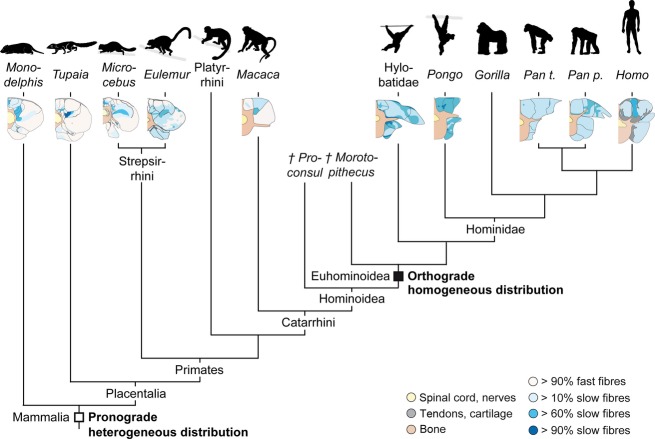
The axial musculoskeletal system is important for the static and dynamic control of the body during both locomotor and non-locomotor behaviour. As a consequence, major evolutionary changes in the positional habits of a species are reflected by morpho-functional adaptations of the axial system. Because of the remarkable phenotypic plasticity of muscle tissue, a close relationship exists between muscle morphology and function. One way to explore major evolutionary transitions in muscle function is therefore by comparative analysis of fibre type composition. In this study, the three-dimensional distribution of slow and fast muscle fibres was analysed in the lumbar perivertebral muscles of two lemuriform (mouse lemur, brown lemur) and four hominoid primate species (white-handed gibbon, orangutan, bonobo, chimpanzee) in order to develop a plausible scenario for the evolution of the contractile properties of the axial muscles in hominoids and to discern possible changes in muscle physiology that were associated with the evolution of orthogrady. Similar to all previously studied quadrupedal mammals, the lemuriform primates in this study exhibited a morpho-functional dichotomy between deep slow contracting local stabilizer muscles and superficial fast contracting global mobilizers and stabilizers and thus retained the fibre distribution pattern typical for quadrupedal non-primates. In contrast, the hominoid primates showed no regionalization of the fibre types, similar to previous observations in Homo. We suggest that this homogeneous fibre composition is associated with the high functional versatility of the axial musculature that was brought about by the evolution of orthograde behaviours and reflects the broad range of mechanical demands acting on the trunk in orthograde hominoids. Because orthogrady is a derived character of euhominoids, the uniform fibre type distribution is hypothesized to coincide with the evolution of orthograde behaviours.
Keywords: autochthonous; epaxial; great ape; hypaxial; lesser ape; skeletal musculature.
© 2013 Anatomical Society.
Figures

Similar articles
-
Functional differentiation of the human lumbar perivertebral musculature revisited by means of muscle fibre type composition.Ann Anat. 2013 Dec;195(6):570-80. doi: 10.1016/j.aanat.2013.07.003. Epub 2013 Aug 20. Ann Anat. 2013. PMID: 24028860
-
Metabolic profile of the perivertebral muscles in small therian mammals: implications for the evolution of the mammalian trunk musculature.Zoology (Jena). 2009;112(4):279-304. doi: 10.1016/j.zool.2008.09.007. Epub 2009 Apr 16. Zoology (Jena). 2009. PMID: 19375292
-
Giant pandas (Carnivora: Ailuropoda melanoleuca) and living hominoids converge on lumbar vertebral adaptations to orthograde trunk posture.J Hum Evol. 2015 Nov;88:160-179. doi: 10.1016/j.jhevol.2015.06.015. Epub 2015 Sep 2. J Hum Evol. 2015. PMID: 26341032
-
Soft-tissue anatomy of the extant hominoids: a review and phylogenetic analysis.J Anat. 2002 Jan;200(Pt 1):3-49. doi: 10.1046/j.0021-8782.2001.00001.x. J Anat. 2002. PMID: 11833653 Free PMC article. Review.
-
On the origin, homologies and evolution of primate facial muscles, with a particular focus on hominoids and a suggested unifying nomenclature for the facial muscles of the Mammalia.J Anat. 2009 Sep;215(3):300-19. doi: 10.1111/j.1469-7580.2009.01111.x. Epub 2009 Jun 12. J Anat. 2009. PMID: 19531159 Free PMC article. Review.
Cited by
-
A 3D musculoskeletal model of the western lowland gorilla hind limb: moment arms and torque of the hip, knee and ankle.J Anat. 2017 Oct;231(4):568-584. doi: 10.1111/joa.12651. Epub 2017 Jul 17. J Anat. 2017. PMID: 28718217 Free PMC article.
-
The evolution of human fatigue resistance.J Comp Physiol B. 2022 Jul;192(3-4):411-422. doi: 10.1007/s00360-022-01439-4. Epub 2022 May 12. J Comp Physiol B. 2022. PMID: 35552490 Free PMC article. Review.
-
Fiber type composition of epaxial muscles is geared toward facilitating rapid spinal extension in the leaper Galago senegalensis.Am J Phys Anthropol. 2018 May;166(1):95-106. doi: 10.1002/ajpa.23405. Epub 2018 Jan 10. Am J Phys Anthropol. 2018. PMID: 29318571 Free PMC article.
-
Substrate oxidation in primary human skeletal muscle cells is influenced by donor age.Cell Tissue Res. 2020 Dec;382(3):599-608. doi: 10.1007/s00441-020-03275-w. Epub 2020 Sep 8. Cell Tissue Res. 2020. PMID: 32897419 Free PMC article.
-
Unique skull network complexity of Tyrannosaurus rex among land vertebrates.Sci Rep. 2019 Feb 6;9(1):1520. doi: 10.1038/s41598-018-37976-8. Sci Rep. 2019. PMID: 30728455 Free PMC article.
References
-
- Acosta L, Roy RR. Fiber-type composition of selected hindlimb muscles of a primate (Cynomolgus monkey. Anat Rec. 1987;218:136–141. - PubMed
-
- Alba DM. Fossil apes from the Vallès-Penedès Basin. Evol Anthropol. 2012;21:254–269. - PubMed
-
- Anapol FC, Jungers WL. Architectural and histochemical diversity within the quadriceps femoris of the brown lemur (Lemur fulvus. Am J Phys Anthropol. 1986;69:355–375. - PubMed
-
- Anapol FC, Jungers WL. Telemetered electromyography of the fast and slow extensors of the leg of the brown lemur (Lemur fulvus. J Exp Biol. 1987;130:341–358. - PubMed
-
- Ariano MA, Armstrong RB, Edgerton VR. Hindlimb muscle fiber populations of five mammals. J Histochem Cytochem. 1973;21:51–55. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
