

Measuring individual locomotor rhythms in honey bees, paper wasps and other similar-sized insects
- PMID: 24436380
- PMCID: PMC3990357
- DOI: 10.1242/jeb.096180
Measuring individual locomotor rhythms in honey bees, paper wasps and other similar-sized insects
Abstract
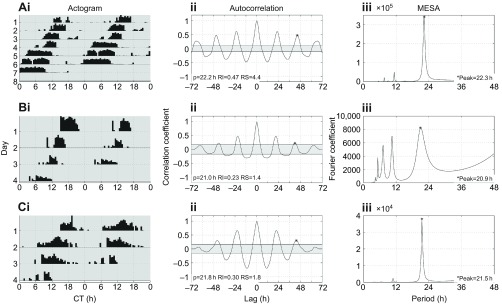
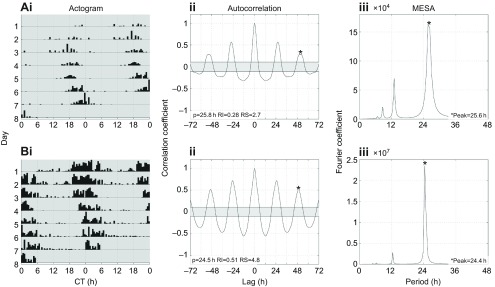
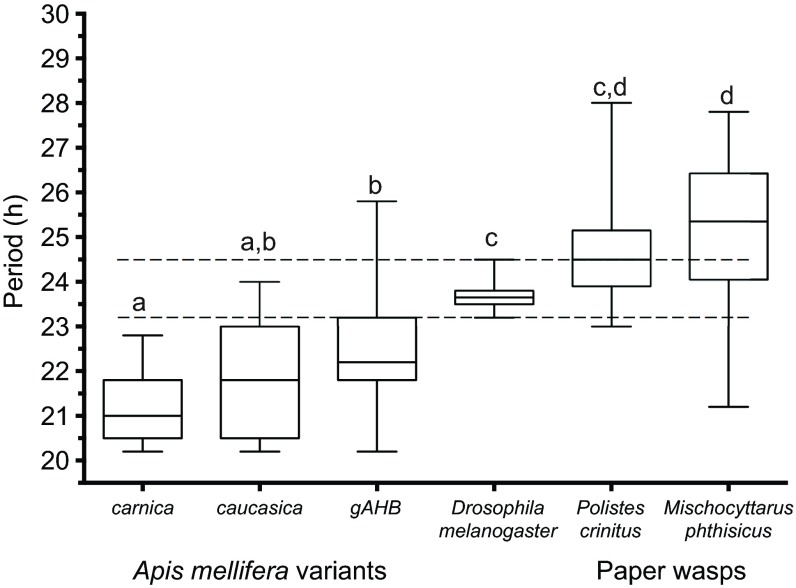
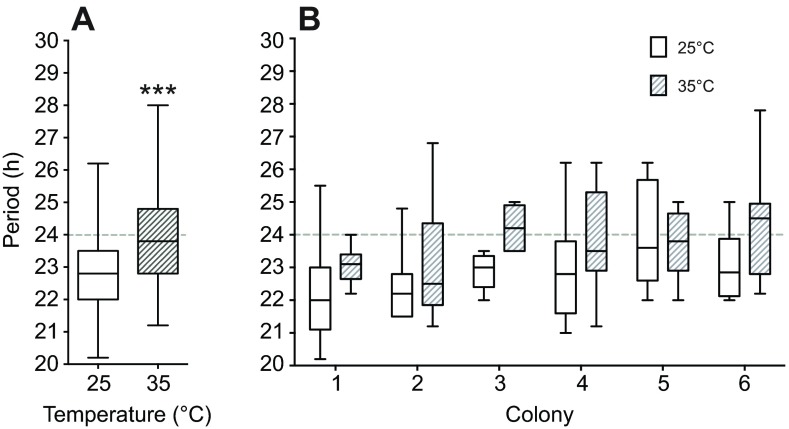
Circadian rhythms in social insects are highly plastic and are modulated by multiple factors. In addition, complex behaviors such as sun-compass orientation and time learning are clearly regulated by the circadian system in these organisms. Despite these unique features of social insect clocks, the mechanisms as well as the functional and evolutionary relevance of these traits remain largely unknown. Here we show a modification of the Drosophila activity monitoring (DAM) system that allowed us to measure locomotor rhythms of the honey bee, Apis mellifera (three variants; gAHB, carnica and caucasica), and two paper wasps (Polistes crinitus and Mischocyttarus phthisicus). A side-by-side comparison of the endogenous period under constant darkness (free-running period) led us to the realization that these social insects exhibit significant deviations from the Earth's 24 h rotational period as well as a large degree of inter-individual variation compared with Drosophila. Experiments at different temperatures, using honey bees as a model, revealed that testing the endogenous rhythm at 35°C, which is the hive's core temperature, results in average periods closer to 24 h compared with 25°C (23.8 h at 35°C versus 22.7 h at 25°C). This finding suggests that the degree of tuning of circadian temperature compensation varies among different organisms. We expect that the commercial availability, cost-effectiveness and integrated nature of this monitoring system will facilitate the growth of the circadian field in these social insects and catalyze our understanding of the mechanisms as well as the functional and evolutionary relevance of circadian rhythms.
Keywords: Apis mellifera; Circadian rhythms; Honey bees; Locomotor activity; Mischocyttarus; Polistes; Temperature compensation; Wasps.
Figures

References
-
- Bahrndorff S., Kjærsgaard A., Pertoldi C., Loeschcke V., Schou T. M., Skovgård H., Hald B. (2012). The effects of sex-ratio and density on locomotor activity in the house fly, Musca domestica. J. Insect Sci. 12, 71
-
- Bertossa R. C., van Dijk J., Beersma D. G., Beukeboom L. W. (2010). Circadian rhythms of adult emergence and activity but not eclosion in males of the parasitic wasp Nasonia vitripennis. J. Insect Physiol. 56, 805-812 - PubMed
-
- Bloch G. (2010). The social clock of the honeybee. J. Biol. Rhythms 25, 307-317 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
