

Structure of cyclin G-associated kinase (GAK) trapped in different conformations using nanobodies
- PMID: 24438162
- PMCID: PMC3957475
- DOI: 10.1042/BJ20131399
Structure of cyclin G-associated kinase (GAK) trapped in different conformations using nanobodies
Abstract
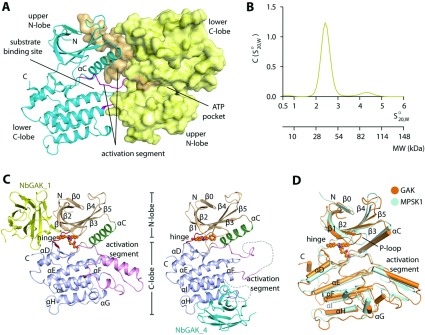
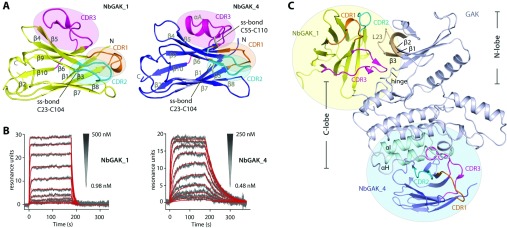
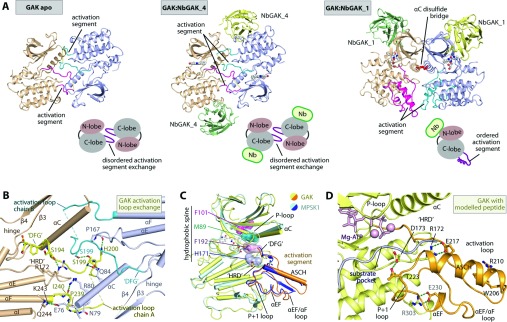
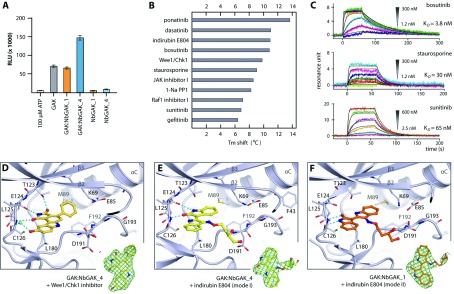
GAK (cyclin G-associated kinase) is a key regulator of clathrin-coated vesicle trafficking and plays a central role during development. Additionally, due to the unusually high plasticity of its catalytic domain, it is a frequent 'off-target' of clinical kinase inhibitors associated with respiratory side effects of these drugs. In the present paper, we determined the crystal structure of the GAK catalytic domain alone and in complex with specific single-chain antibodies (nanobodies). GAK is constitutively active and weakly associates in solution. The GAK apo structure revealed a dimeric inactive state of the catalytic domain mediated by an unusual activation segment interaction. Co-crystallization with the nanobody NbGAK_4 trapped GAK in a dimeric arrangement similar to the one observed in the apo structure, whereas NbGAK_1 captured the activation segment of monomeric GAK in a well-ordered conformation, representing features of the active kinase. The presented structural and biochemical data provide insight into the domain plasticity of GAK and demonstrate the utility of nanobodies to gain insight into conformational changes of dynamic molecules. In addition, we present structural data on the binding mode of ATP mimetic inhibitors and enzyme kinetic data, which will support rational inhibitor design of inhibitors to reduce the off-target effect on GAK.
Figures
Similar articles
-
Role of cyclin G-associated kinase in uncoating clathrin-coated vesicles from non-neuronal cells.J Biol Chem. 2000 Jan 14;275(2):1365-70. doi: 10.1074/jbc.275.2.1365. J Biol Chem. 2000. PMID: 10625686
-
Canonical interaction of cyclin G associated kinase with adaptor protein 1 regulates lysosomal enzyme sorting.Mol Biol Cell. 2007 Aug;18(8):2991-3001. doi: 10.1091/mbc.e06-12-1162. Epub 2007 May 30. Mol Biol Cell. 2007. PMID: 17538018 Free PMC article.
-
GAK, a regulator of clathrin-mediated membrane trafficking, localizes not only in the cytoplasm but also in the nucleus.Genes Cells. 2009 May;14(5):627-41. doi: 10.1111/j.1365-2443.2009.01296.x. Epub 2009 Apr 15. Genes Cells. 2009. PMID: 19371378
-
Nanobodies to Study G Protein-Coupled Receptor Structure and Function.Annu Rev Pharmacol Toxicol. 2017 Jan 6;57:19-37. doi: 10.1146/annurev-pharmtox-010716-104710. Epub 2016 Dec 7. Annu Rev Pharmacol Toxicol. 2017. PMID: 27959623 Free PMC article. Review.
-
Quantitative assessment of the association between GAK rs1564282 C/T polymorphism and the risk of Parkinson's disease.J Clin Neurosci. 2015 Jul;22(7):1077-80. doi: 10.1016/j.jocn.2014.12.014. Epub 2015 May 11. J Clin Neurosci. 2015. PMID: 25975492 Review.
Cited by
-
Micro RNAs-The Small Big Players in Hepatitis E Virus Infection: A Comprehensive Review.Biomolecules. 2022 Oct 22;12(11):1543. doi: 10.3390/biom12111543. Biomolecules. 2022. PMID: 36358893 Free PMC article. Review.
-
Nanobodies as Probes for Protein Dynamics in Vitro and in Cells.J Biol Chem. 2016 Feb 19;291(8):3767-75. doi: 10.1074/jbc.R115.679811. Epub 2015 Dec 16. J Biol Chem. 2016. PMID: 26677230 Free PMC article. Review.
-
A designed ankyrin-repeat protein that targets Parkinson's disease-associated LRRK2.J Biol Chem. 2024 Jul;300(7):107469. doi: 10.1016/j.jbc.2024.107469. Epub 2024 Jun 12. J Biol Chem. 2024. PMID: 38876305 Free PMC article.
-
An improved yeast surface display platform for the screening of nanobody immune libraries.Sci Rep. 2019 Jan 23;9(1):382. doi: 10.1038/s41598-018-37212-3. Sci Rep. 2019. PMID: 30674983 Free PMC article.
-
From discovery to treatment: tracing the path of hepatitis E virus.Virol J. 2024 Aug 23;21(1):194. doi: 10.1186/s12985-024-02470-3. Virol J. 2024. PMID: 39180020 Free PMC article. Review.
References
-
- Manning G., Whyte D. B., Martinez R., Hunter T., Sudarsanam S. The protein kinase complement of the human genome. Science. 2002;298:1912–1934. - PubMed
-
- Kanaoka Y., Kimura S. H., Okazaki I., Ikeda M., Nojima H. GAK: a cyclin G associated kinase contains a tensin/auxilin-like domain. FEBS Lett. 1997;402:73–80. - PubMed
-
- Lee D. W., Zhao X., Zhang F., Eisenberg E., Greene L. E. Depletion of GAK/auxilin 2 inhibits receptor-mediated endocytosis and recruitment of both clathrin and clathrin adaptors. J. Cell Sci. 2005;118:4311–4321. - PubMed
-
- Zhang C. X., Engqvist-Goldstein A. E., Carreno S., Owen D. J., Smythe E., Drubin D. G. Multiple roles for cyclin G-associated kinase in clathrin-mediated sorting events. Traffic. 2005;6:1103–1113. - PubMed
-
- Shimizu H., Nagamori I., Yabuta N., Nojima H. GAK, a regulator of clathrin-mediated membrane traffic, also controls centrosome integrity and chromosome congression. J. Cell Sci. 2009;122:3145–3152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases