

Signaling through the adaptor molecule MyD88 in CD4+ T cells is required to overcome suppression by regulatory T cells
- PMID: 24439266
- PMCID: PMC4445716
- DOI: 10.1016/j.immuni.2013.10.023
Signaling through the adaptor molecule MyD88 in CD4+ T cells is required to overcome suppression by regulatory T cells
Erratum in
- Immunity. 2014 May 15;40(5):814
Abstract
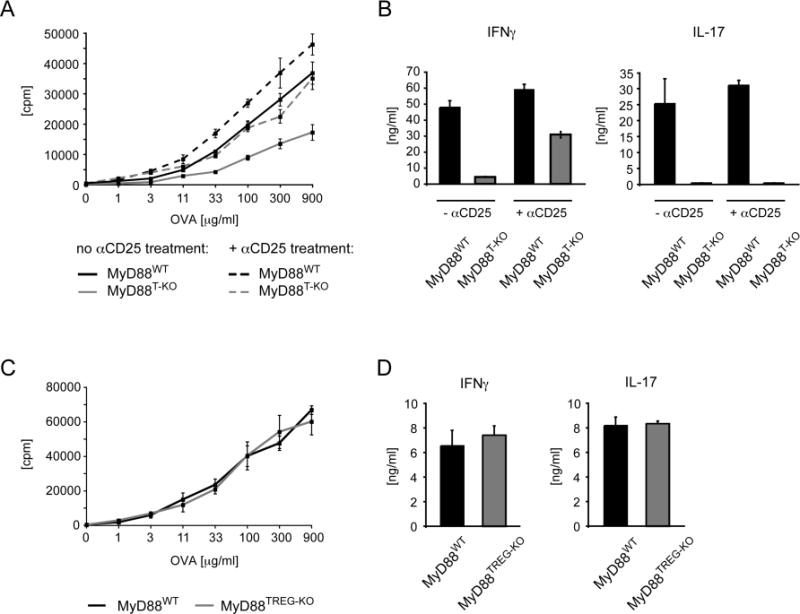
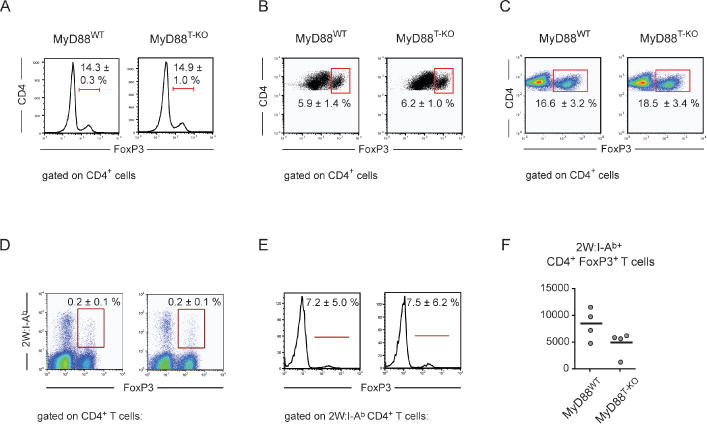
Innate immune recognition controls adaptive immune responses through multiple mechanisms. The MyD88 signaling adaptor operates in many cell types downstream of Toll-like receptors (TLRs) and interleukin-1 (IL-1) receptor family members. Cell-type-specific functions of MyD88 signaling remain poorly characterized. Here, we have shown that the T cell-specific ablation of MyD88 in mice impairs not only T helper 17 (Th17) cell responses, but also Th1 cell responses. MyD88 relayed signals of TLR-induced IL-1, which became dispensable for Th1 cell responses in the absence of T regulatory (Treg) cells. Treg cell-specific ablation of MyD88 had no effect, suggesting that IL-1 acts on naive CD4(+) T cells instead of Treg cells themselves. Together, these findings demonstrate that IL-1 renders naive CD4(+) T cells refractory to Treg cell-mediated suppression in order to allow their differentiation into Th1 cells. In addition, IL-1 was also important for the generation of functional CD4(+) memory T cells.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
T cell-intrinsic role of IL-6 signaling in primary and memory responses.Elife. 2014 May 19;3:e01949. doi: 10.7554/eLife.01949. Elife. 2014. PMID: 24842874 Free PMC article.
-
MyD88 is essential to sustain mTOR activation necessary to promote T helper 17 cell proliferation by linking IL-1 and IL-23 signaling.Proc Natl Acad Sci U S A. 2013 Feb 5;110(6):2270-5. doi: 10.1073/pnas.1206048110. Epub 2013 Jan 22. Proc Natl Acad Sci U S A. 2013. PMID: 23341605 Free PMC article.
-
Postoperative ileus involves interleukin-1 receptor signaling in enteric glia.Gastroenterology. 2014 Jan;146(1):176-87.e1. doi: 10.1053/j.gastro.2013.09.030. Epub 2013 Sep 22. Gastroenterology. 2014. PMID: 24067878
-
MyDths and un-TOLLed truths: sensor, instructive and effector immunity to tuberculosis.Immunol Lett. 2008 Feb 15;116(1):15-23. doi: 10.1016/j.imlet.2007.11.015. Epub 2007 Dec 26. Immunol Lett. 2008. PMID: 18191460 Review.
-
TIR8/SIGIRR: an IL-1R/TLR family member with regulatory functions in inflammation and T cell polarization.Trends Immunol. 2009 Sep;30(9):439-46. doi: 10.1016/j.it.2009.06.001. Epub 2009 Aug 21. Trends Immunol. 2009. PMID: 19699681 Review.
Cited by
-
Toll-Like Receptor 2 Mediates In Vivo Pro- and Anti-inflammatory Effects of Mycobacterium Tuberculosis and Modulates Autoimmune Encephalomyelitis.Front Immunol. 2016 May 24;7:191. doi: 10.3389/fimmu.2016.00191. eCollection 2016. Front Immunol. 2016. PMID: 27252700 Free PMC article.
-
Evaluation of the Effects of Genistein In Vitro as a Chemopreventive Agent for Colorectal Cancer-Strategy to Improve Its Efficiency When Administered Orally.Molecules. 2022 Oct 19;27(20):7042. doi: 10.3390/molecules27207042. Molecules. 2022. PMID: 36296636 Free PMC article.
-
T cell-intrinsic role of IL-6 signaling in primary and memory responses.Elife. 2014 May 19;3:e01949. doi: 10.7554/eLife.01949. Elife. 2014. PMID: 24842874 Free PMC article.
-
Induction and activation of human Th17 by targeting antigens to dendritic cells via dectin-1.J Immunol. 2014 Jun 15;192(12):5776-88. doi: 10.4049/jimmunol.1301661. Epub 2014 May 16. J Immunol. 2014. PMID: 24835401 Free PMC article. Clinical Trial.
-
Overproduction of IL-2 by Cbl-b deficient CD4+ T cells provides resistance against regulatory T cells.Oncoimmunology. 2020 Apr 7;9(1):1737368. doi: 10.1080/2162402X.2020.1737368. eCollection 2020. Oncoimmunology. 2020. PMID: 32313719 Free PMC article.
References
-
- Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A, Sallusto F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat Immunol. 2007;8:942–949. - PubMed
-
- Ben Ahmed M, Belhadj Hmida N, Moes N, Buyse S, Abdeladhim M, Louzir H, Cerf-Bensussan N. IL-15 renders conventional lymphocytes resistant to suppressive functions of regulatory T cells through activation of the phosphatidylinositol 3-kinase pathway. J Immunol. 2009;182:6763–6770. - PubMed
-
- Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. - PubMed
-
- Brodsky IE, Monack D. NLR-mediated control of inflammasome assembly in the host response against bacterial pathogens. Semin Immunol. 2009;21:199–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
