

Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation
- PMID: 24439267
- PMCID: PMC3923301
- DOI: 10.1016/j.immuni.2013.11.019
Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation
Abstract
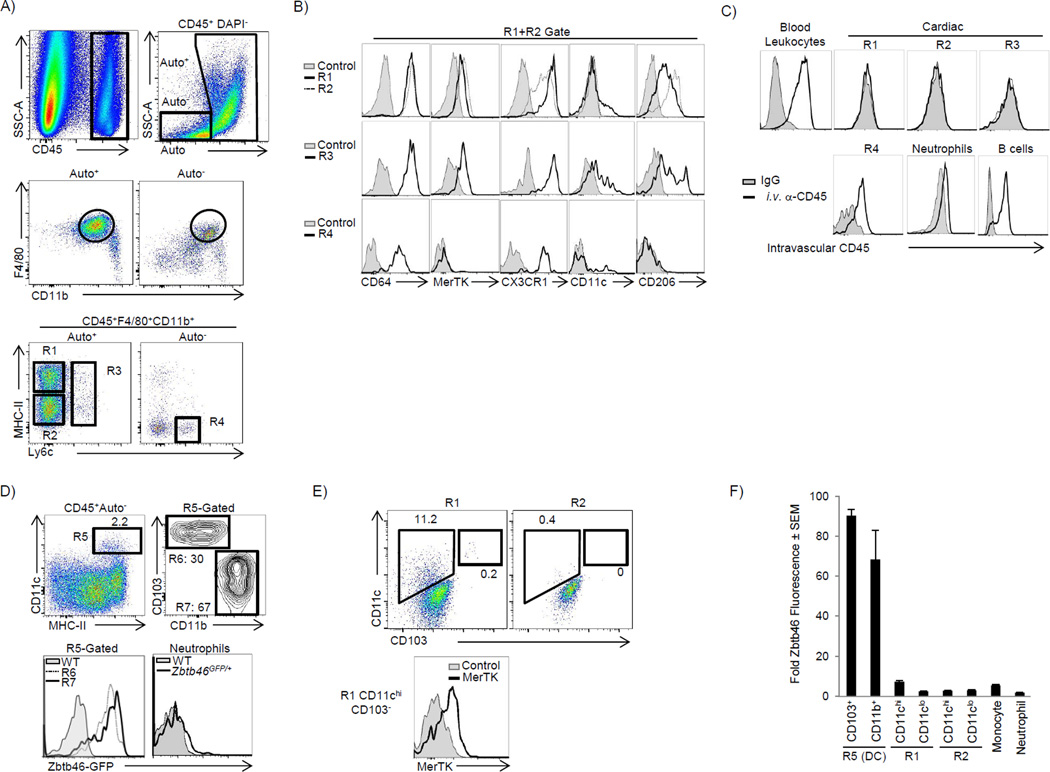
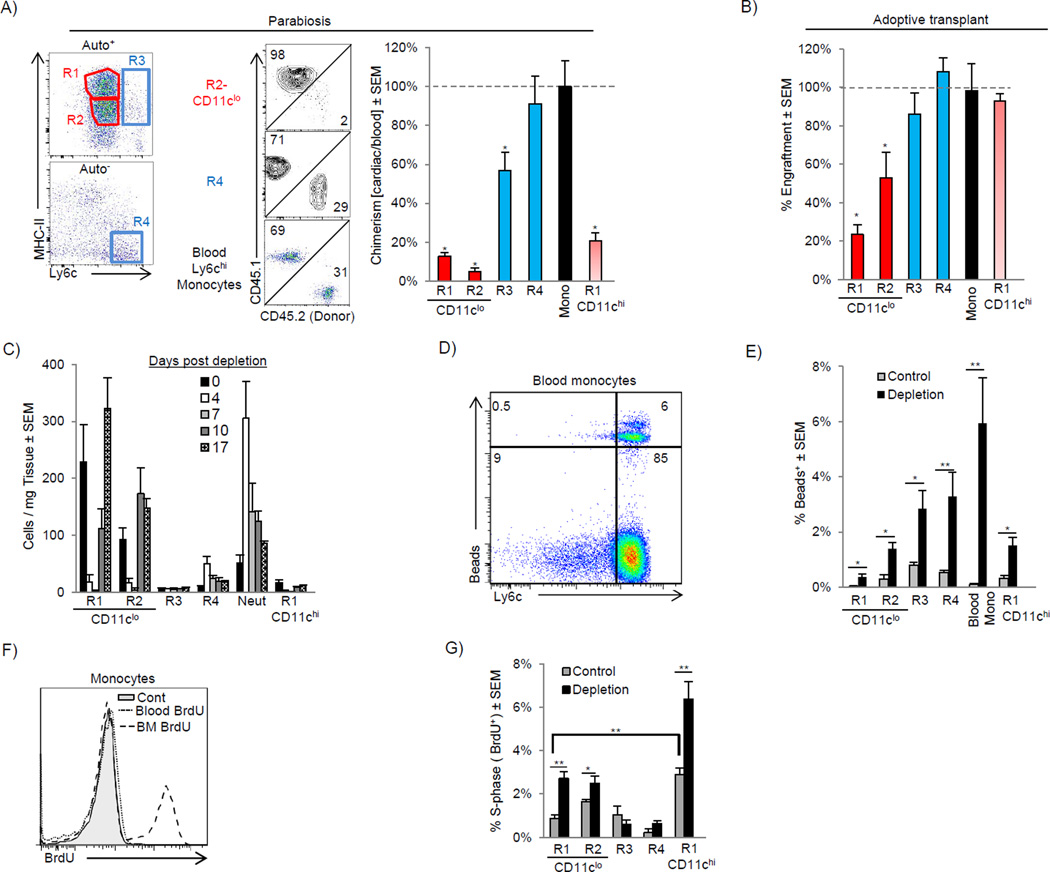
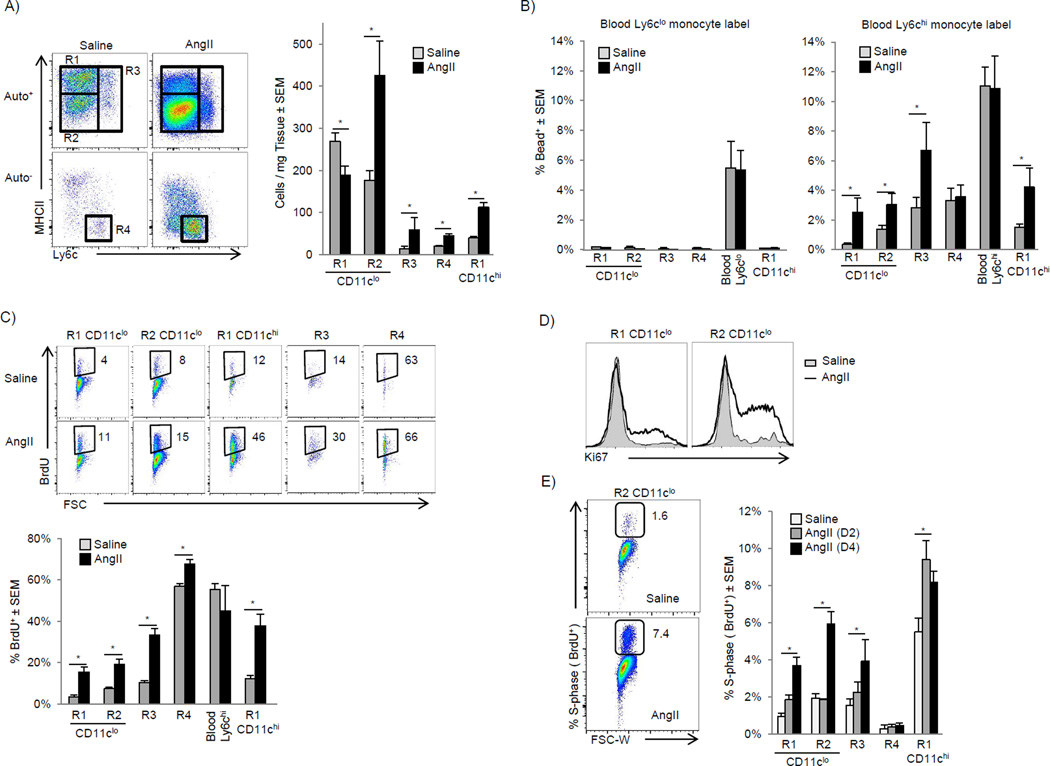
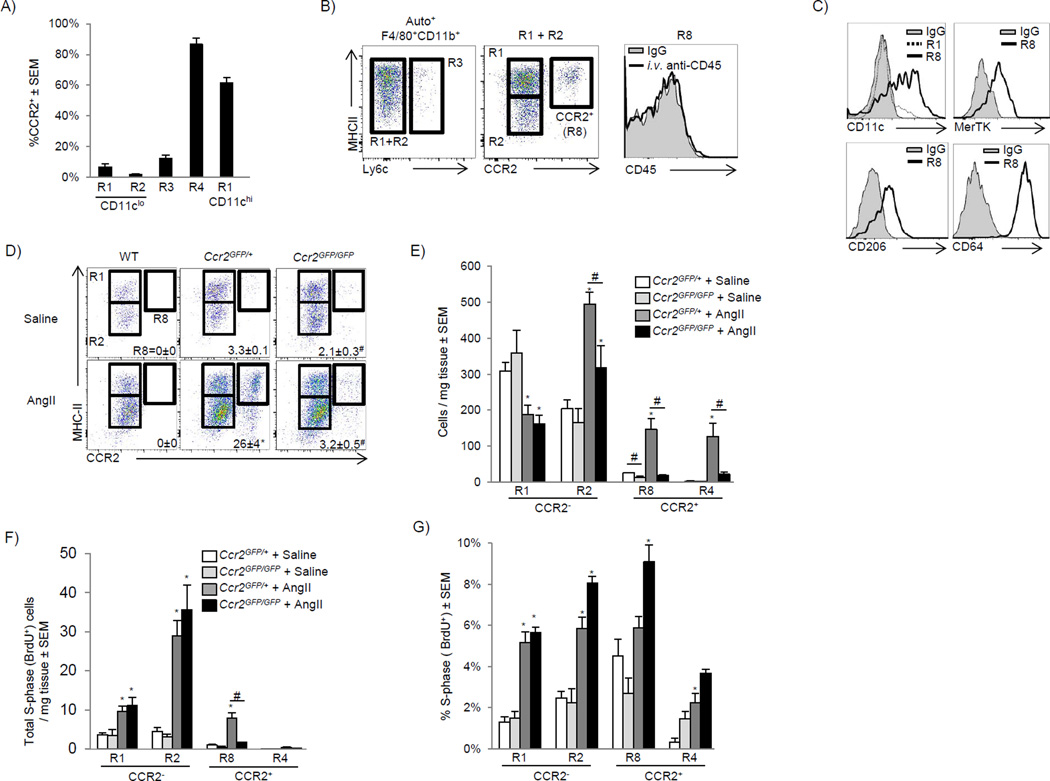
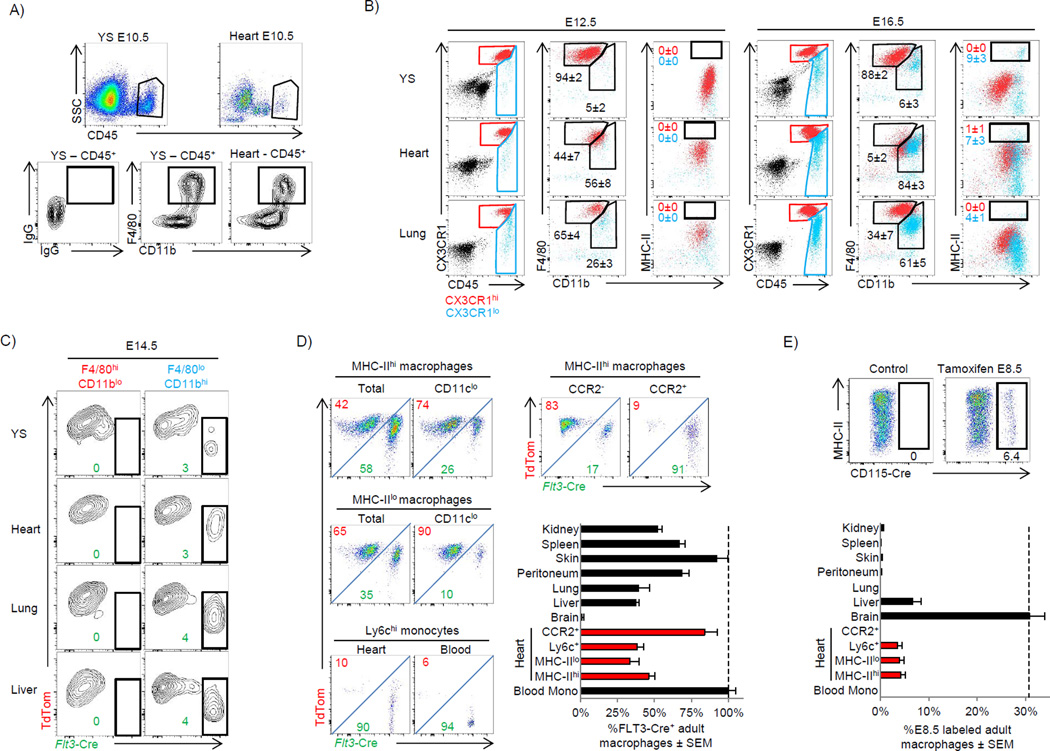
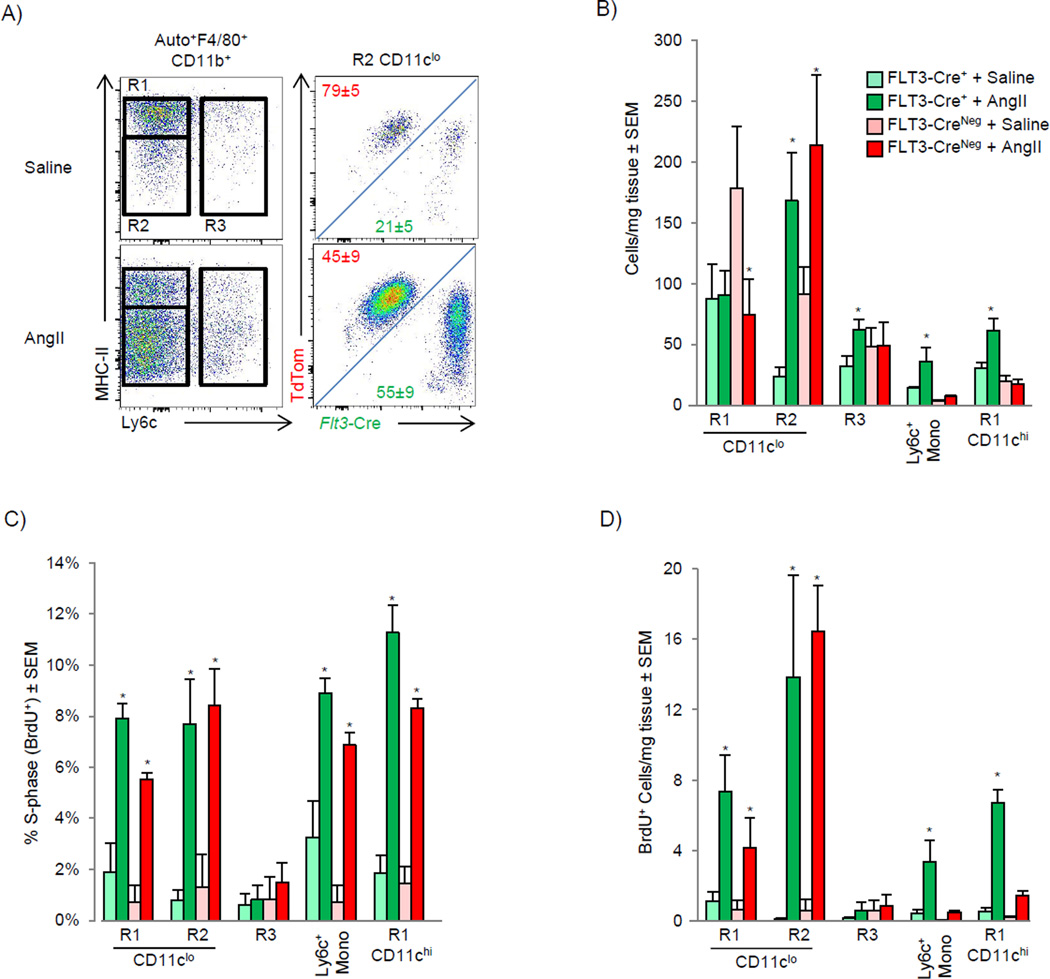
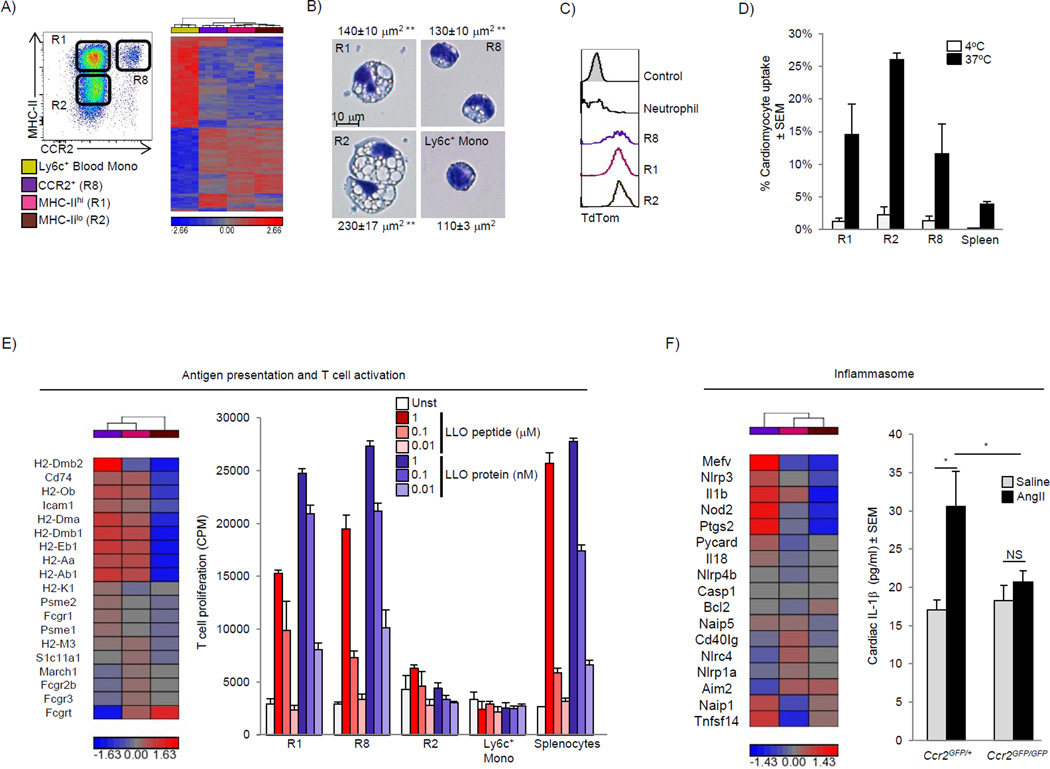
Cardiac macrophages are crucial for tissue repair after cardiac injury but are not well characterized. Here we identify four populations of cardiac macrophages. At steady state, resident macrophages were primarily maintained through local proliferation. However, after macrophage depletion or during cardiac inflammation, Ly6c(hi) monocytes contributed to all four macrophage populations, whereas resident macrophages also expanded numerically through proliferation. Genetic fate mapping revealed that yolk-sac and fetal monocyte progenitors gave rise to the majority of cardiac macrophages, and the heart was among a minority of organs in which substantial numbers of yolk-sac macrophages persisted in adulthood. CCR2 expression and dependence distinguished cardiac macrophages of adult monocyte versus embryonic origin. Transcriptional and functional data revealed that monocyte-derived macrophages coordinate cardiac inflammation, while playing redundant but lesser roles in antigen sampling and efferocytosis. These data highlight the presence of multiple cardiac macrophage subsets, with different functions, origins, and strategies to regulate compartment size.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Cardiac macrophages: how to mend a broken heart.Immunity. 2014 Jan 16;40(1):3-5. doi: 10.1016/j.immuni.2013.12.005. Immunity. 2014. PMID: 24439261
References
-
- Francis GS. Neurohormonal control of heart failure. Cleve. Clin. J. Med. 2011;78(Suppl):S75–S79. - PubMed
-
- Frangogiannis NG, Dewald O, Xia Y, Ren G, Haudek S, Leucker T, Kraemer D, Taffet G, Rollins BJ, Entman ML. Critical role of monocyte chemoattractant protein-1/CC chemokine ligand 2 in the pathogenesis of ischemic cardiomyopathy. Circulation. 2007;115:584–592. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
