

Local F-actin network links synapse formation and axon branching
- PMID: 24439377
- PMCID: PMC3954643
- DOI: 10.1016/j.cell.2013.12.009
Local F-actin network links synapse formation and axon branching
Abstract
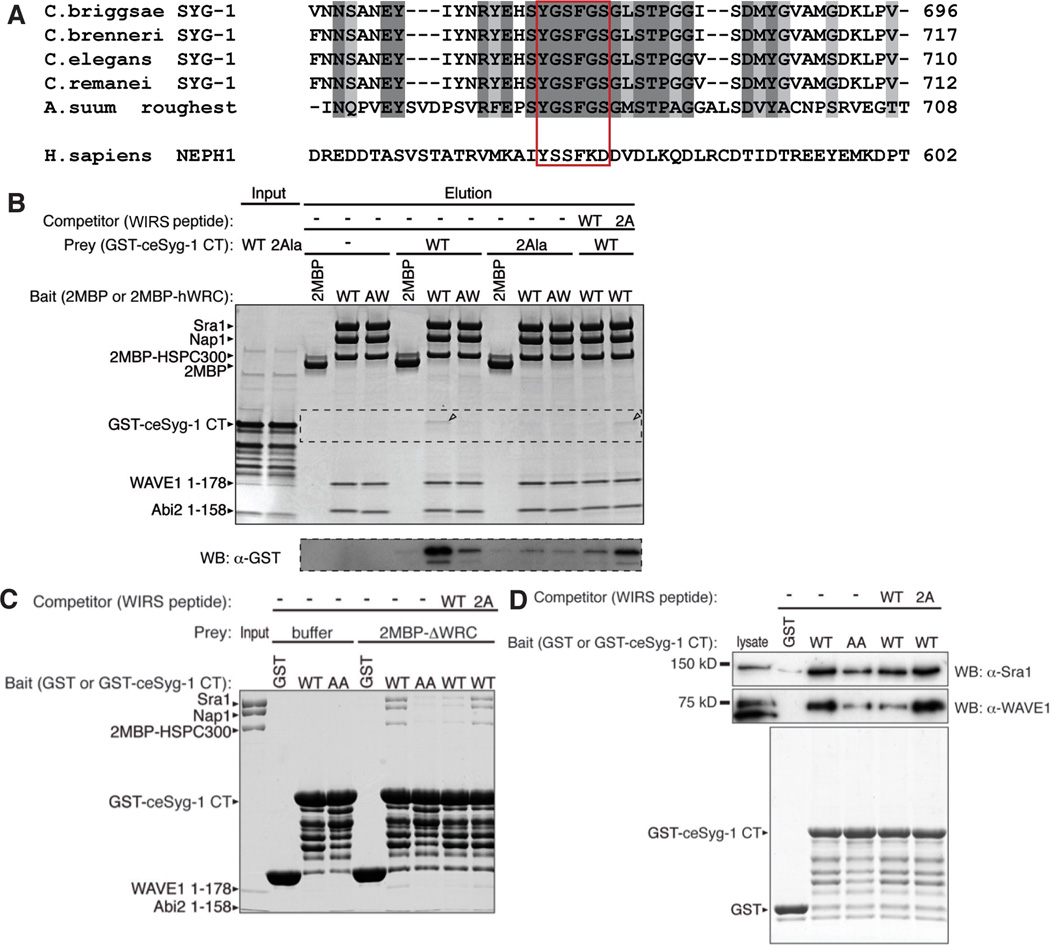
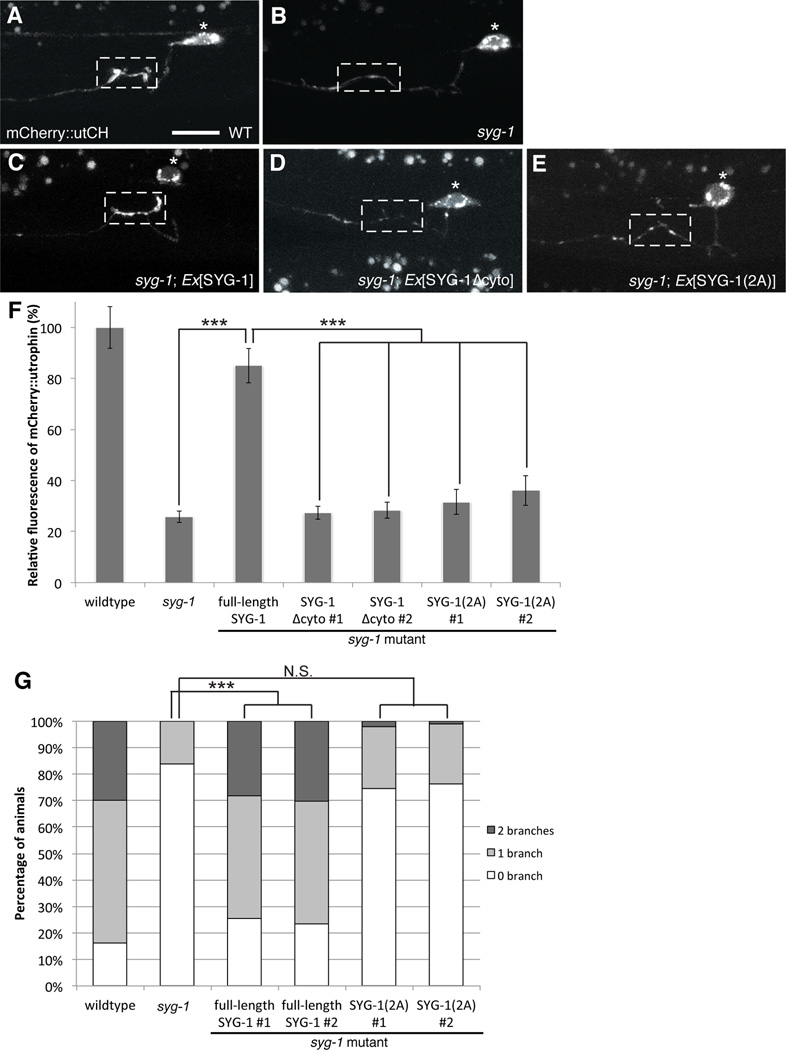
Axonal branching and synapse formation are tightly linked developmental events during the establishment of synaptic circuits. Newly formed synapses promote branch initiation and stability. However, little is known about molecular mechanisms that link these two processes. Here, we show that local assembly of an F-actin cytoskeleton at nascent presynaptic sites initiates both synapse formation and axon branching. We further find that assembly of the F-actin network requires a direct interaction between the synaptic cell adhesion molecule SYG-1 and a key regulator of actin cytoskeleton, the WVE-1/WAVE regulatory complex (WRC). SYG-1 cytoplasmic tail binds to the WRC using a consensus WRC interacting receptor sequence (WIRS). WRC mutants or mutating the SYG-1 WIRS motif leads to loss of local F-actin, synaptic material, and axonal branches. Together, these data suggest that synaptic adhesion molecules, which serve as a necessary component for both synaptogenesis and axonal branch formation, directly regulate subcellular actin cytoskeletal organization.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
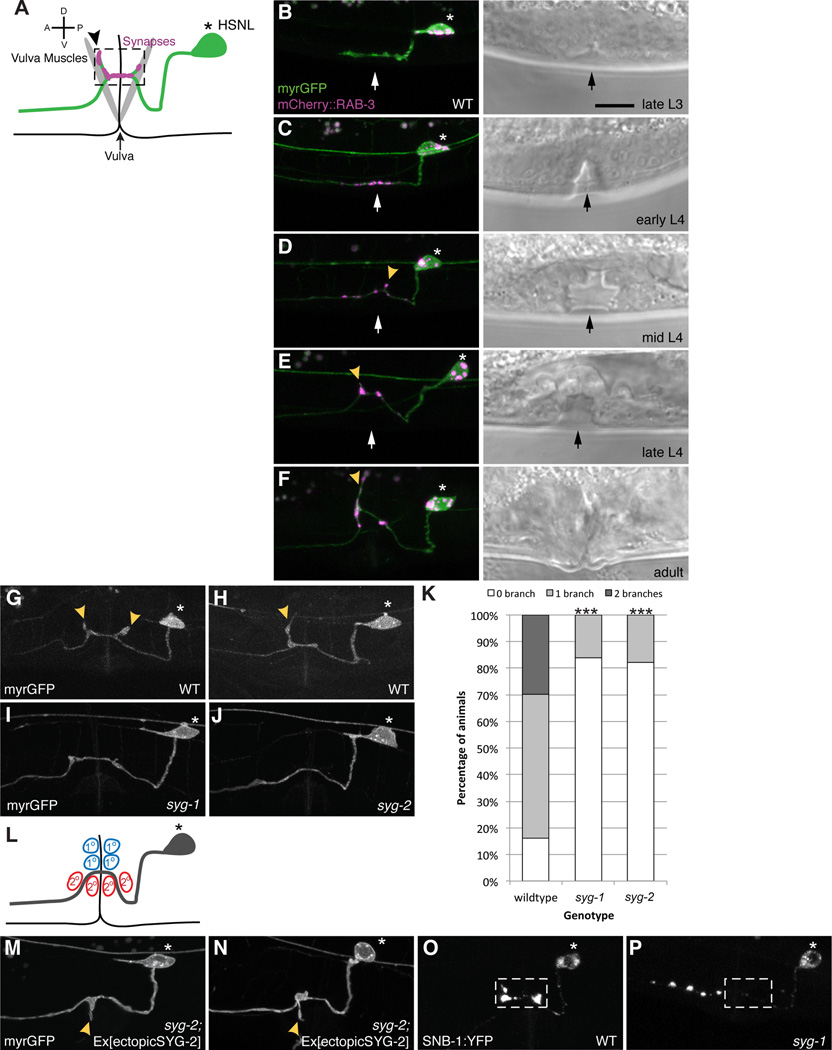
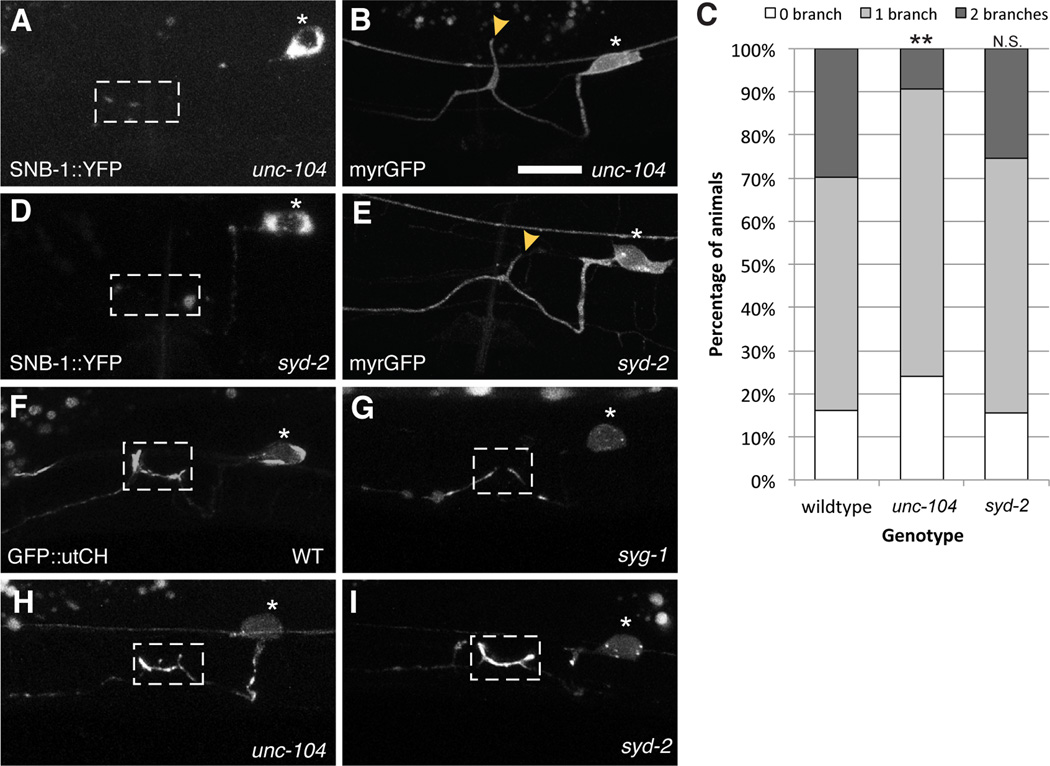
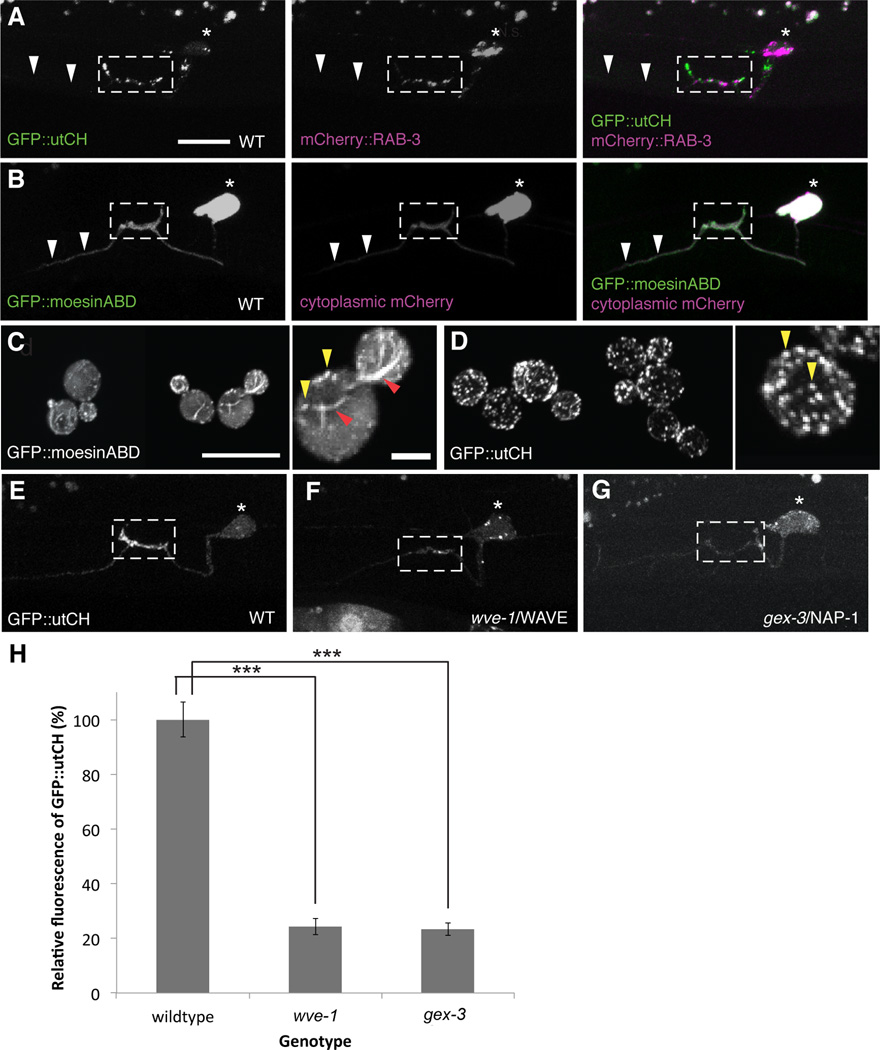
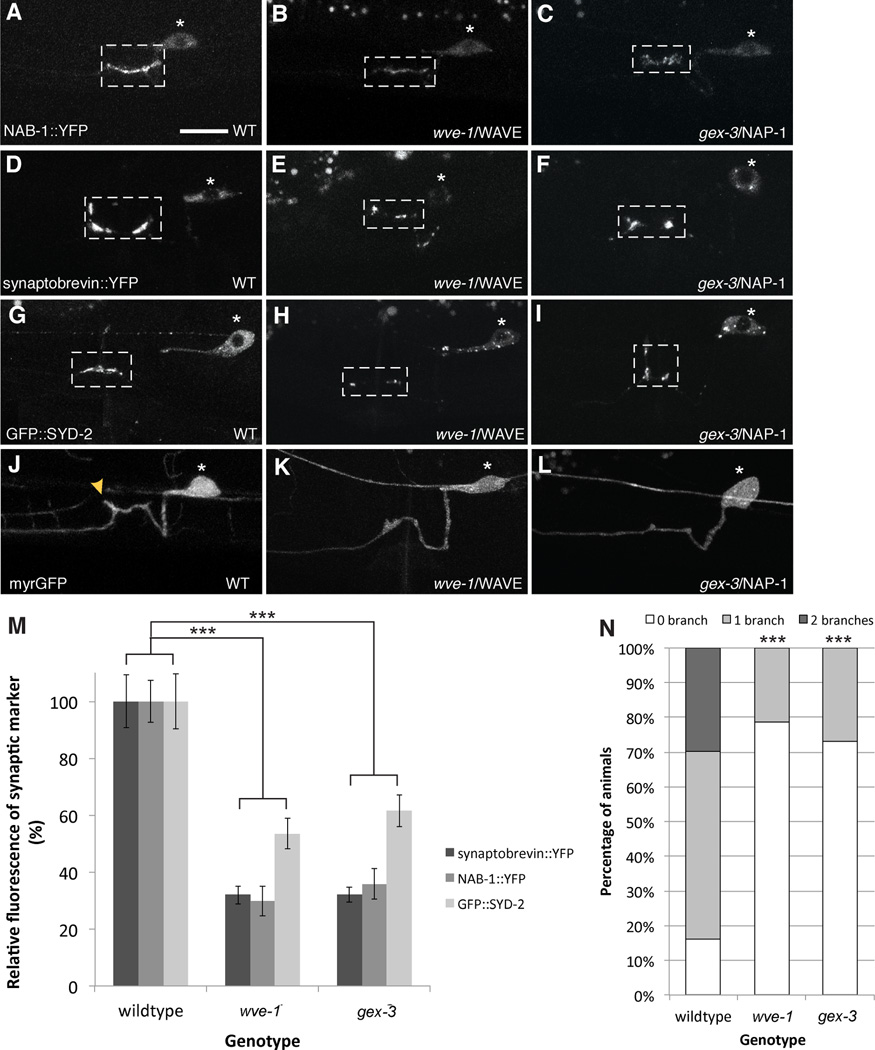
Comment in
-
The shape of things to come.Cell. 2014 Jan 16;156(1-2):13-4. doi: 10.1016/j.cell.2013.12.037. Cell. 2014. PMID: 24439365
References
-
- Chen SX, Tari PK, She K, Haas K. Neurexin-Neuroligin Cell Adhesion Complexes Contribute to Synaptotropic Dendritogenesis via Growth Stabilization Mechanisms In Vivo. Neuron. 2010a;67:967–983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
