

The SMC condensin complex is required for origin segregation in Bacillus subtilis
- PMID: 24440393
- PMCID: PMC3947903
- DOI: 10.1016/j.cub.2013.11.050
The SMC condensin complex is required for origin segregation in Bacillus subtilis
Abstract
SMC condensin complexes play a central role in organizing and compacting chromosomes in all domains of life [1, 2]. In the bacterium Bacillus subtilis, cells lacking SMC are viable only during slow growth and display decondensed chromosomes, suggesting that SMC complexes function throughout the genome [3, 4]. Here, we show that rapid inactivation of SMC or its partner protein ScpB during fast growth leads to a failure to resolve newly replicated origins and a complete block to chromosome segregation. Importantly, the loss of origin segregation is not due to an inability to unlink precatenated sister chromosomes by Topoisomerase IV. In support of the idea that ParB-mediated recruitment of SMC complexes to the origin is important for their segregation, cells with reduced levels of SMC that lack ParB are severely impaired in origin resolution. Finally, we demonstrate that origin segregation is a task shared by the condensin complex and the parABS partitioning system. We propose that origin-localized SMC constrains adjacent DNA segments along their lengths, drawing replicated origins in on themselves and away from each other. This SMC-mediated lengthwise condensation, bolstered by the parABS system, drives origin segregation.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
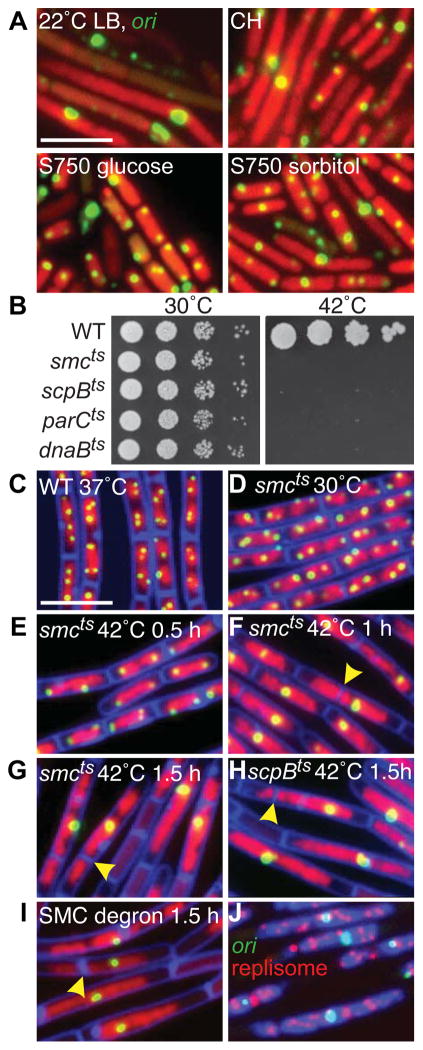
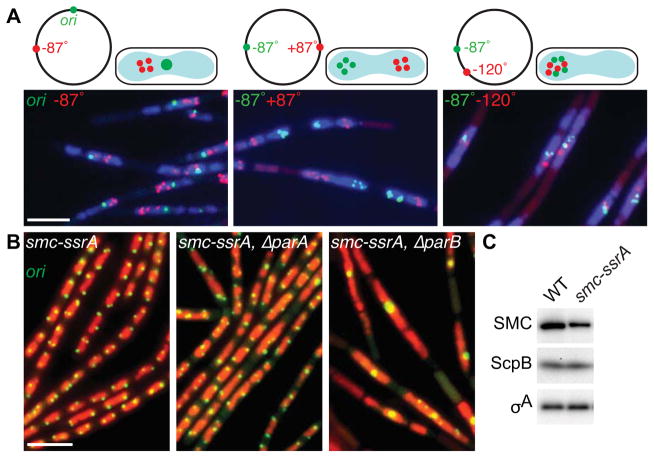
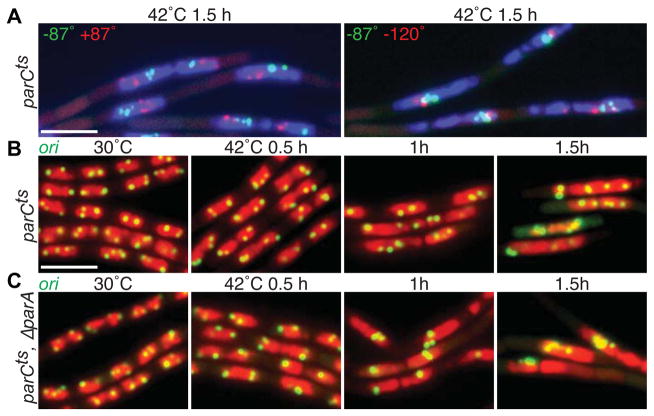
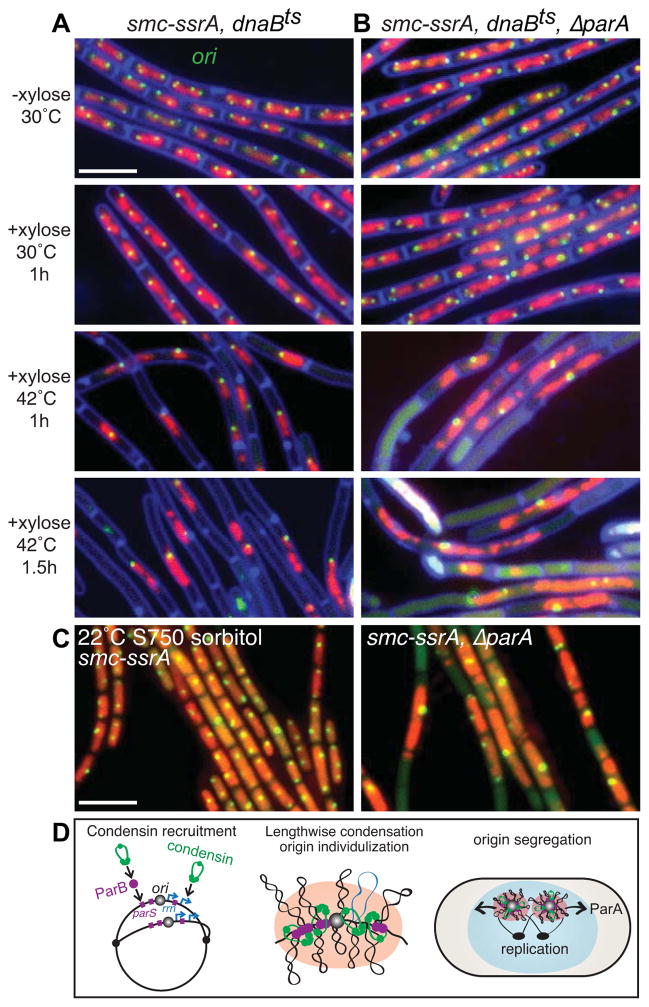
Comment in
-
Chromosome organization: original condensins.Curr Biol. 2014 Feb 3;24(3):R111-3. doi: 10.1016/j.cub.2013.12.033. Curr Biol. 2014. PMID: 24502782
References
-
- Hirano T. At the heart of the chromosome: SMC proteins in action. Nat Rev Mol Cell Biol. 2006;7:311–322. - PubMed
-
- Nasmyth K, Haering CH. The structure and function of SMC and kleisin complexes. Annu Rev Biochem. 2005;74:595–648. - PubMed
-
- Moriya S, Tsujikawa E, Hassan AK, Asai K, Kodama T, Ogasawara N. A Bacillus subtilis gene-encoding protein homologous to eukaryotic SMC motor protein is necessary for chromosome partition. Mol Microbiol. 1998;29:179–187. - PubMed
-
- Cui Y, Petrushenko ZM, Rybenkov VV. MukB acts as a macromolecular clamp in DNA condensation. Nat Struct Mol Biol. 2008;15:411–418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
