

ZFHX4 interacts with the NuRD core member CHD4 and regulates the glioblastoma tumor-initiating cell state
- PMID: 24440720
- PMCID: PMC4041390
- DOI: 10.1016/j.celrep.2013.12.032
ZFHX4 interacts with the NuRD core member CHD4 and regulates the glioblastoma tumor-initiating cell state
Abstract
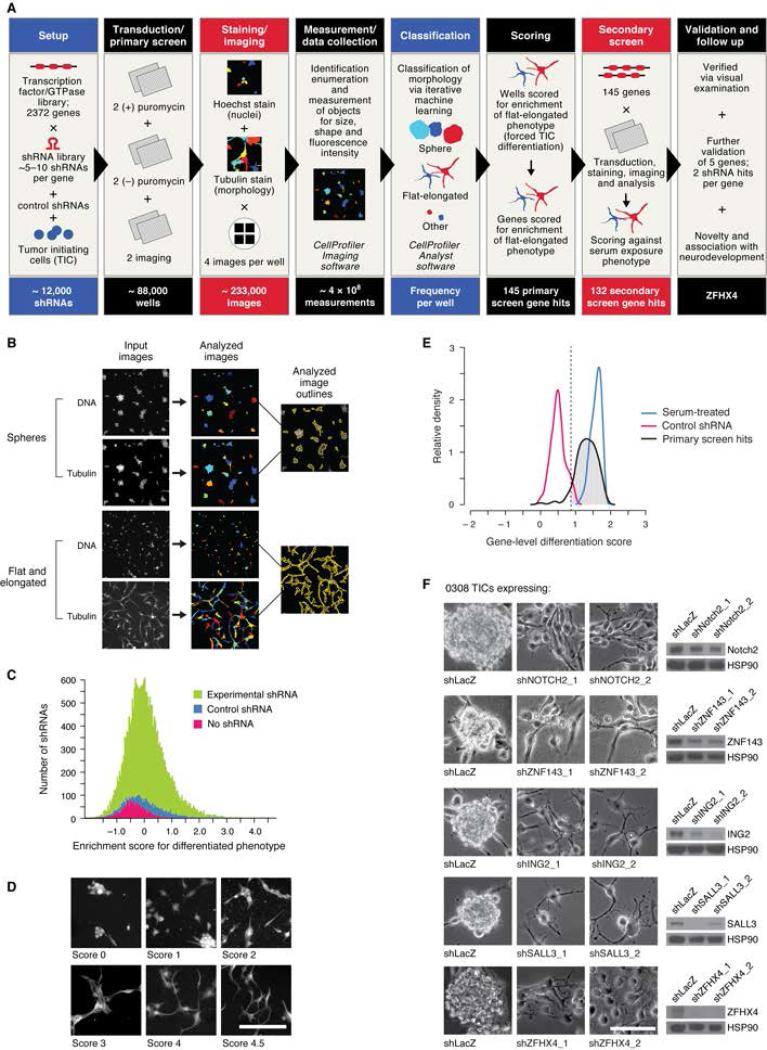
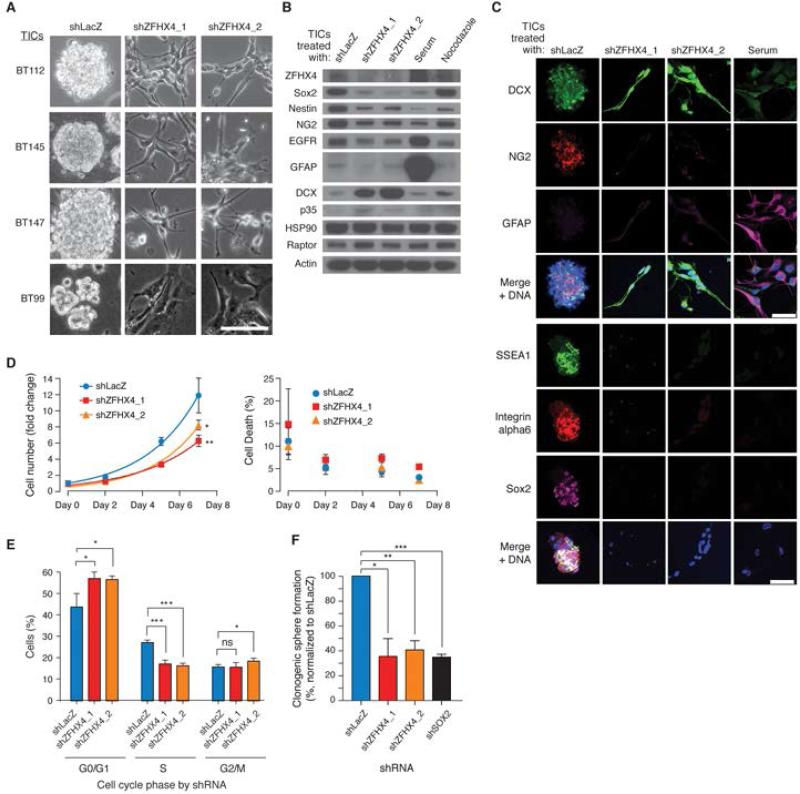
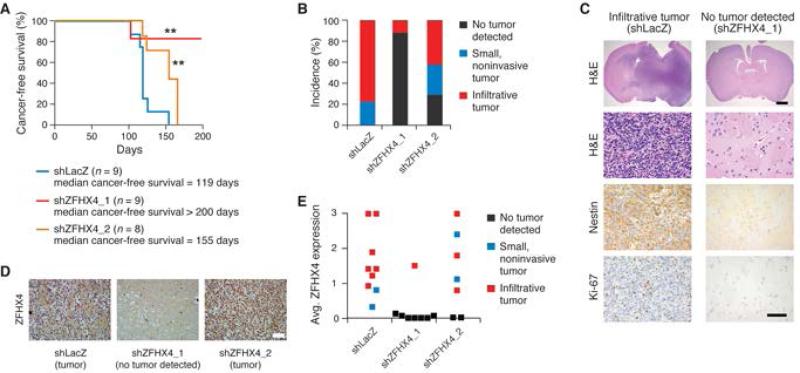
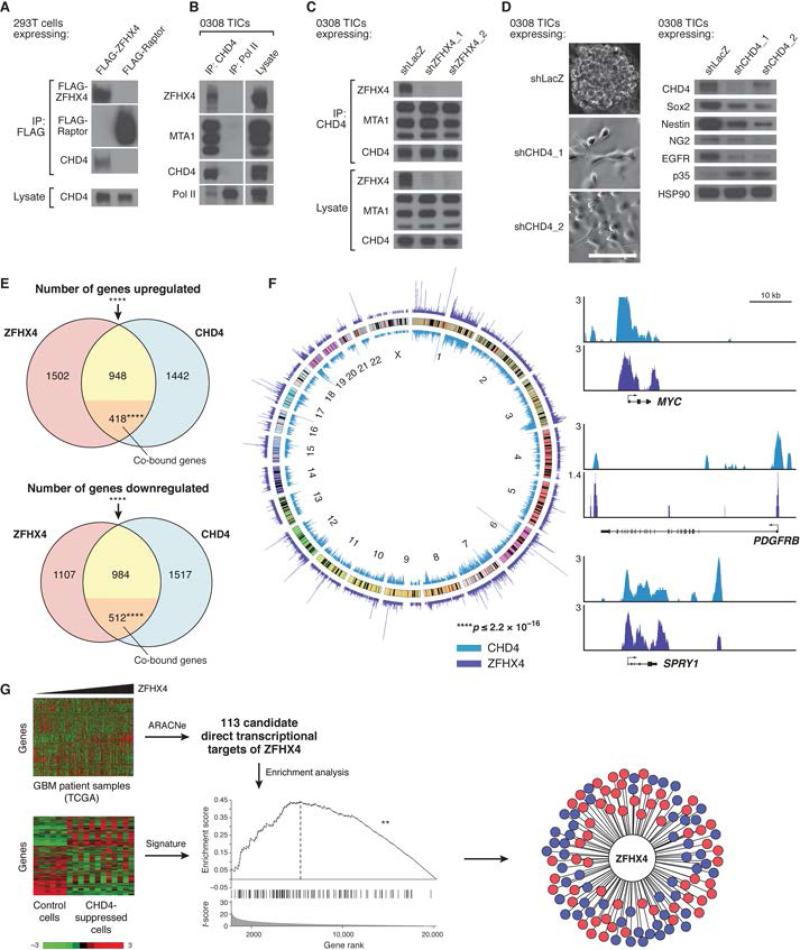
Glioblastoma (GBM) harbors subpopulations of therapy-resistant tumor-initiating cells (TICs) that are self-renewing and multipotent. To understand the regulation of the TIC state, we performed an image-based screen for genes regulating GBM TIC maintenance and identified ZFHX4, a 397 kDa transcription factor. ZFHX4 is required to maintain TIC-associated and normal human neural precursor cell phenotypes in vitro, suggesting that ZFHX4 regulates differentiation, and its suppression increases glioma-free survival in intracranial xenografts. ZFHX4 interacts with CHD4, a core member of the nucleosome remodeling and deacetylase (NuRD) complex. ZFHX4 and CHD4 bind to overlapping sets of genomic loci and control similar gene expression programs. Using expression data derived from GBM patients, we found that ZFHX4 significantly affects CHD4-mediated gene expression perturbations, which defines ZFHX4 as a master regulator of CHD4. These observations define ZFHX4 as a regulatory factor that links the chromatin-remodeling NuRD complex and the GBM TIC state.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, Dewhirst MW, Bigner DD, Rich JN. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444:756–760. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA103866/CA/NCI NIH HHS/United States
- R01NS080944/NS/NINDS NIH HHS/United States
- P30 CA016672/CA/NCI NIH HHS/United States
- R01CA129105/CA/NCI NIH HHS/United States
- U01CA176058/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 CA142536/CA/NCI NIH HHS/United States
- R01CA170592/CA/NCI NIH HHS/United States
- R01CA146455/CA/NCI NIH HHS/United States
- K08NS062907/NS/NINDS NIH HHS/United States
- U01 CA176058/CA/NCI NIH HHS/United States
- P01CA095616/CA/NCI NIH HHS/United States
- K12CA090354/CA/NCI NIH HHS/United States
- R01 CA129105/CA/NCI NIH HHS/United States
- U24 CA143883/CA/NCI NIH HHS/United States
- R01 HG002668/HG/NHGRI NIH HHS/United States
- U54CA121852/CA/NCI NIH HHS/United States
- K08 NS062907/NS/NINDS NIH HHS/United States
- R01HG002668/HG/NHGRI NIH HHS/United States
- K99 AG045144/AG/NIA NIH HHS/United States
- U01 CA168426/CA/NCI NIH HHS/United States
- P01 CA095616/CA/NCI NIH HHS/United States
- P30CA016672/CA/NCI NIH HHS/United States
- R01GM089652/GM/NIGMS NIH HHS/United States
- P01CA142536/CA/NCI NIH HHS/United States
- K12 CA090354/CA/NCI NIH HHS/United States
- R01 NS080944/NS/NINDS NIH HHS/United States
- U01CA168426/CA/NCI NIH HHS/United States
- U24CA143883/CA/NCI NIH HHS/United States
- R01 GM089652/GM/NIGMS NIH HHS/United States
- U54 CA121852/CA/NCI NIH HHS/United States
- R01 CA170592/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
