

A dynamic alternative splicing program regulates gene expression during terminal erythropoiesis
- PMID: 24442673
- PMCID: PMC3973340
- DOI: 10.1093/nar/gkt1388
A dynamic alternative splicing program regulates gene expression during terminal erythropoiesis
Abstract
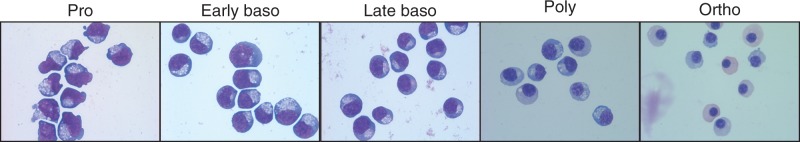
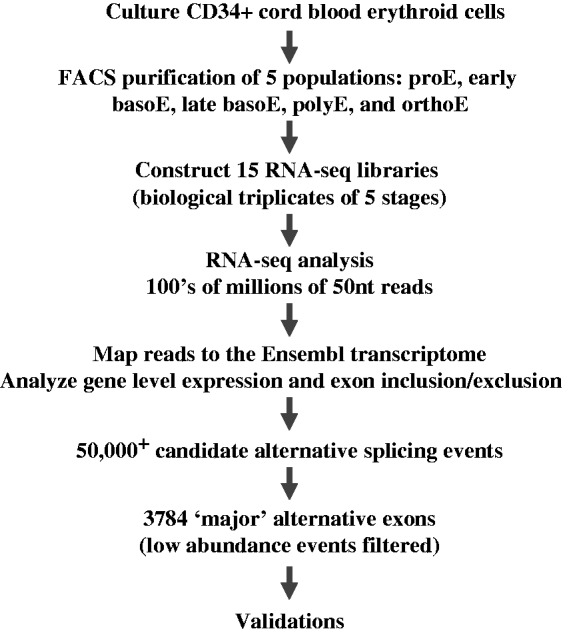
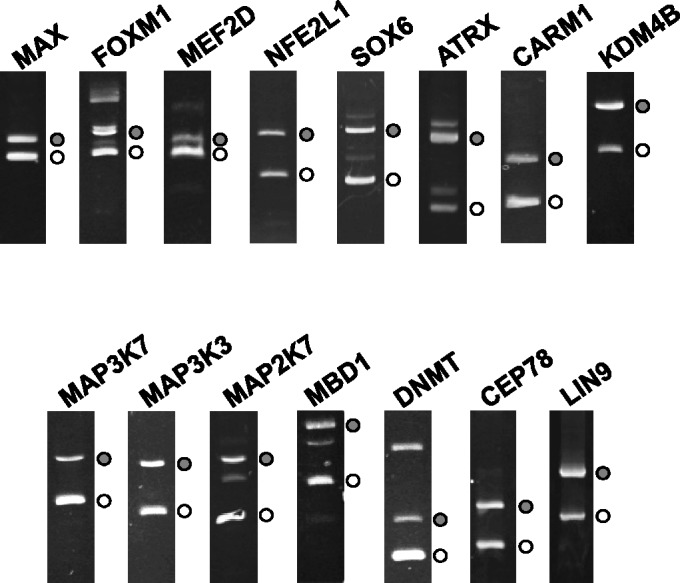
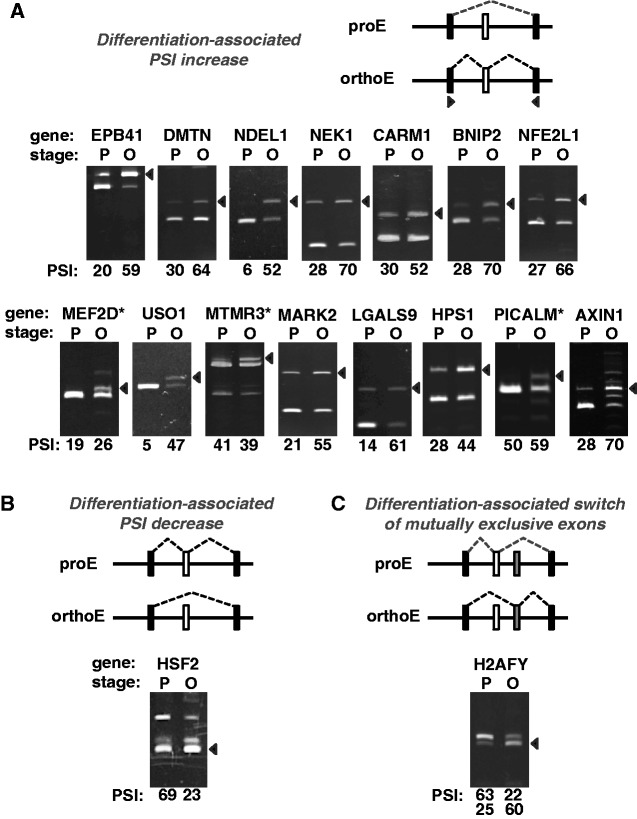
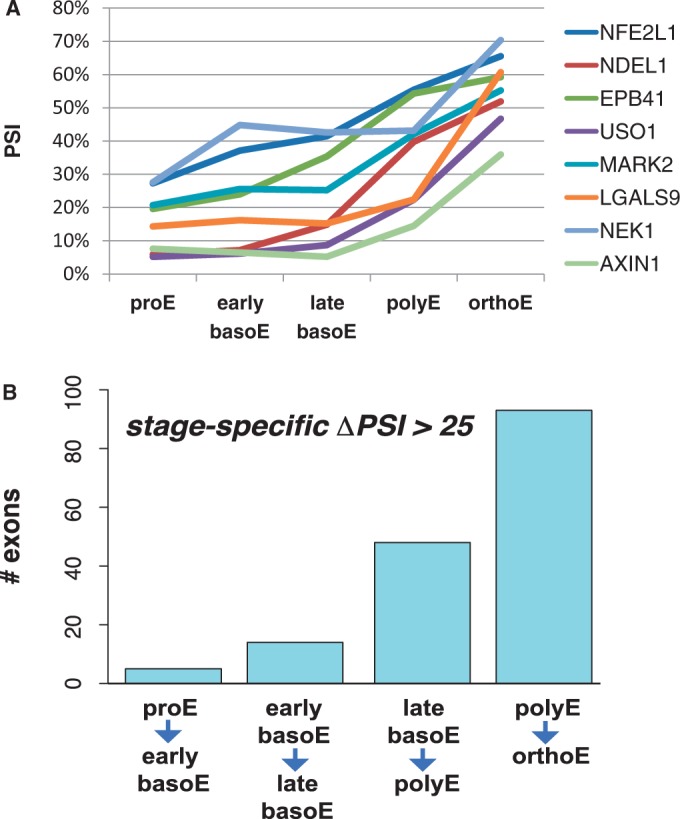
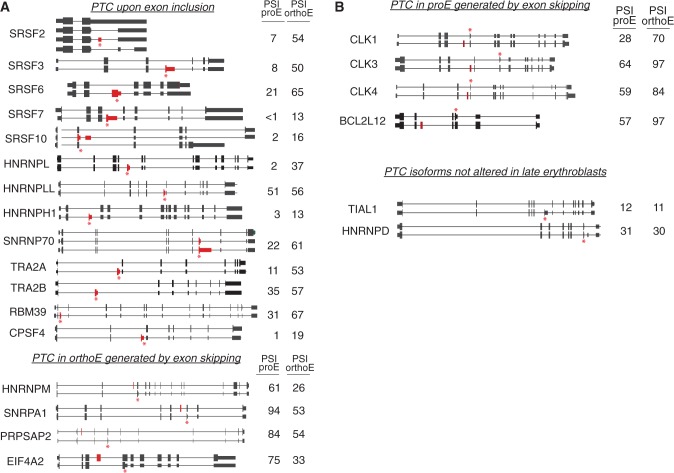
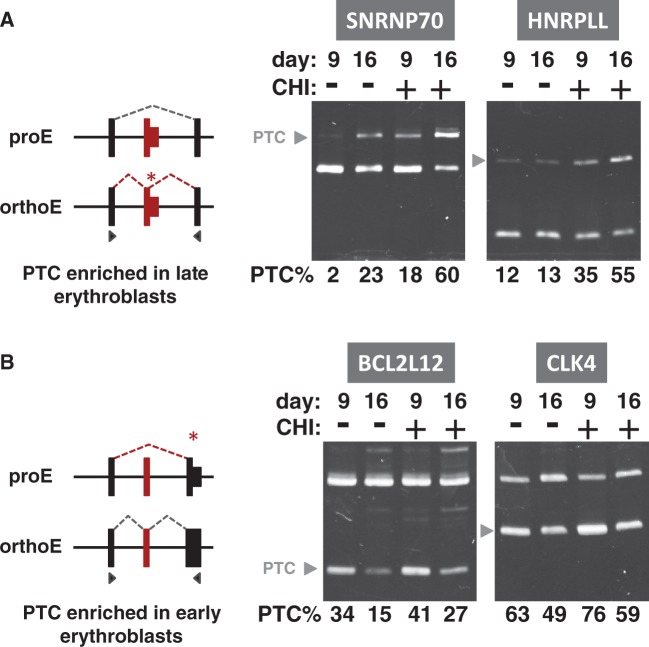
Alternative pre-messenger RNA splicing remodels the human transcriptome in a spatiotemporal manner during normal development and differentiation. Here we explored the landscape of transcript diversity in the erythroid lineage by RNA-seq analysis of five highly purified populations of morphologically distinct human erythroblasts, representing the last four cell divisions before enucleation. In this unique differentiation system, we found evidence of an extensive and dynamic alternative splicing program encompassing genes with many diverse functions. Alternative splicing was particularly enriched in genes controlling cell cycle, organelle organization, chromatin function and RNA processing. Many alternative exons exhibited differentiation-associated switches in splicing efficiency, mostly in late-stage polychromatophilic and orthochromatophilic erythroblasts, in concert with extensive cellular remodeling that precedes enucleation. A subset of alternative splicing switches introduces premature translation termination codons into selected transcripts in a differentiation stage-specific manner, supporting the hypothesis that alternative splicing-coupled nonsense-mediated decay contributes to regulation of erythroid-expressed genes as a novel part of the overall differentiation program. We conclude that a highly dynamic alternative splicing program in terminally differentiating erythroblasts plays a major role in regulating gene expression to ensure synthesis of appropriate proteome at each stage as the cells remodel in preparation for production of mature red cells.
Figures

References
-
- Yoshida K, Sanada M, Shiraishi Y, Nowak D, Nagata Y, Yamamoto R, Sato Y, Sato-Otsubo A, Kon A, Nagasaki M, et al. Frequent pathway mutations of splicing machinery in myelodysplasia. Nature. 2011;478:64–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
