

Calcineurin controls hyphal growth, virulence, and drug tolerance of Candida tropicalis
- PMID: 24442892
- PMCID: PMC4135728
- DOI: 10.1128/EC.00302-13
Calcineurin controls hyphal growth, virulence, and drug tolerance of Candida tropicalis
Abstract
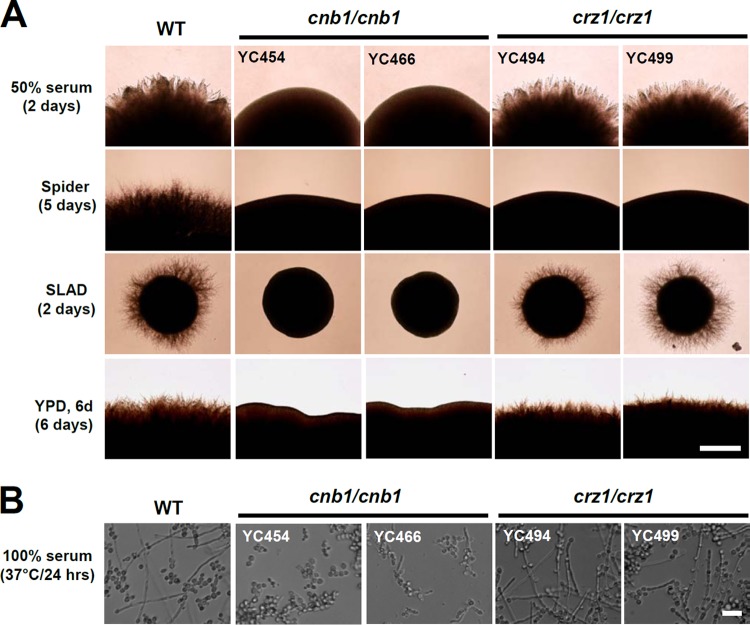
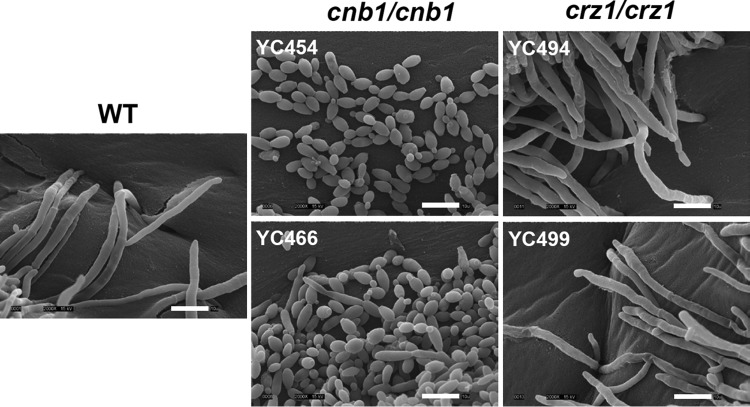
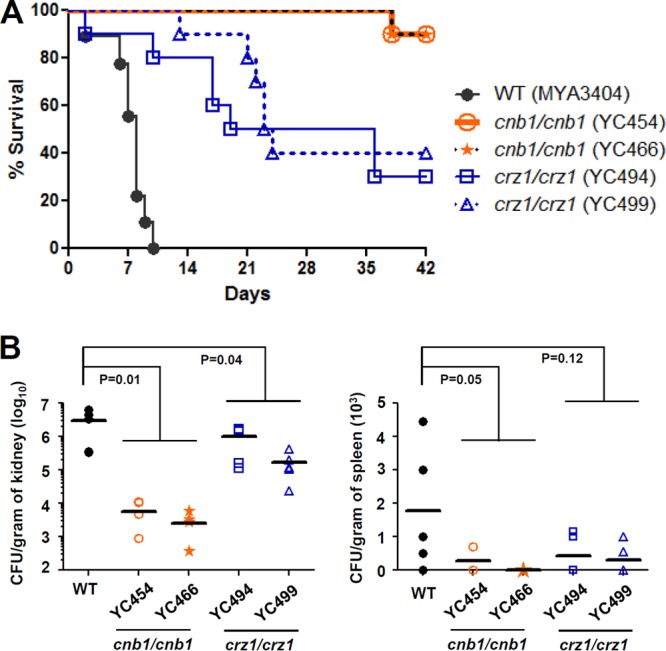
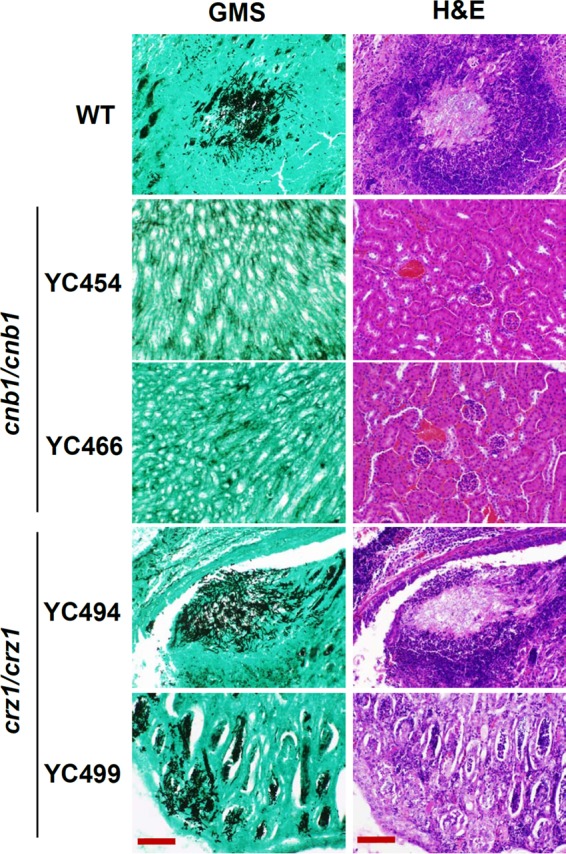
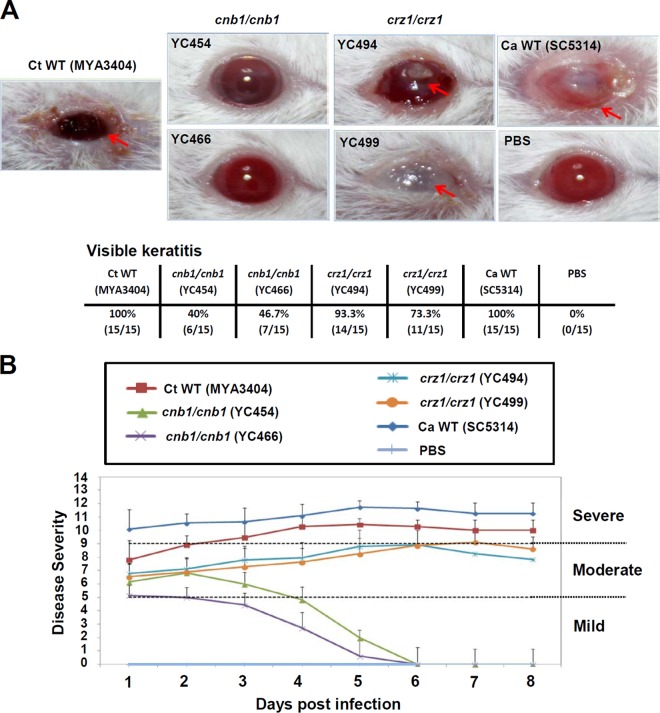
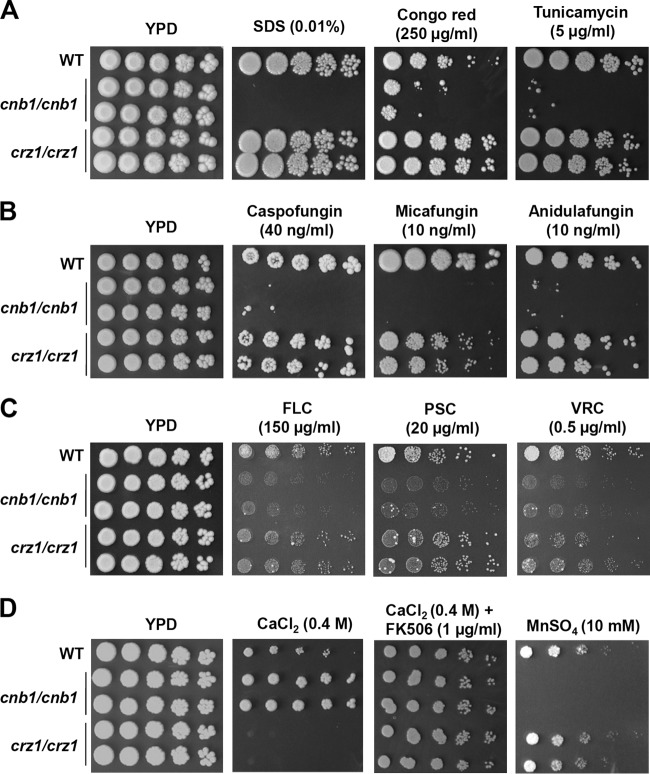
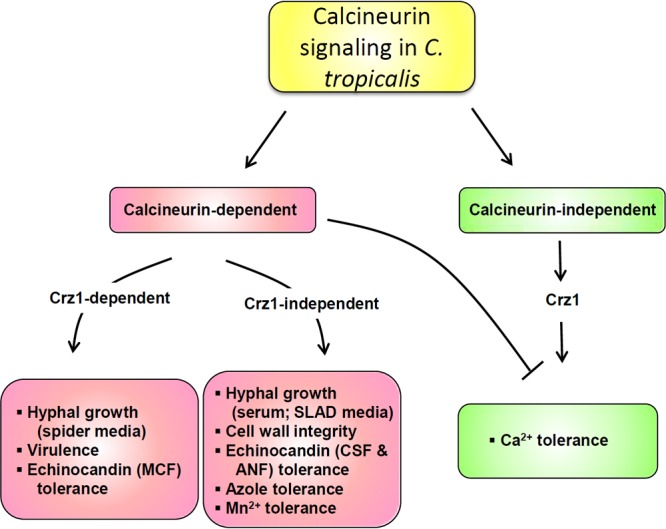
Candida tropicalis, a species closely related to Candida albicans, is an emerging fungal pathogen associated with high mortality rates of 40 to 70%. Like C. albicans and Candida dubliniensis, C. tropicalis is able to form germ tubes, pseudohyphae, and hyphae, but the genes involved in hyphal growth machinery and virulence remain unclear in C. tropicalis. Recently, echinocandin- and azole-resistant C. tropicalis isolates have frequently been isolated from various patients around the world, making treatment difficult. However, studies of the C. tropicalis genes involved in drug tolerance are limited. Here, we investigated the roles of calcineurin and its potential target, Crz1, for core stress responses and pathogenesis in C. tropicalis. We demonstrate that calcineurin and Crz1 are required for hyphal growth, micafungin tolerance, and virulence in a murine systemic infection model, while calcineurin but not Crz1 is essential for tolerance of azoles, caspofungin, anidulafungin, and cell wall-perturbing agents, suggesting that calcineurin has both Crz1-dependent and -independent functions in C. tropicalis. In addition, we found that calcineurin and Crz1 have opposite roles in controlling calcium tolerance. Calcineurin serves as a negative regulator, while Crz1 plays a positive role for calcium tolerance in C. tropicalis.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Calcineurin controls drug tolerance, hyphal growth, and virulence in Candida dubliniensis.Eukaryot Cell. 2011 Jun;10(6):803-19. doi: 10.1128/EC.00310-10. Epub 2011 Apr 29. Eukaryot Cell. 2011. PMID: 21531874 Free PMC article.
-
Calcineurin is required for pseudohyphal growth, virulence, and drug resistance in Candida lusitaniae.PLoS One. 2012;7(8):e44192. doi: 10.1371/journal.pone.0044192. Epub 2012 Aug 31. PLoS One. 2012. PMID: 22952924 Free PMC article.
-
Hsp90 governs echinocandin resistance in the pathogenic yeast Candida albicans via calcineurin.PLoS Pathog. 2009 Jul;5(7):e1000532. doi: 10.1371/journal.ppat.1000532. Epub 2009 Jul 31. PLoS Pathog. 2009. PMID: 19649312 Free PMC article.
-
Calcineurin signaling: lessons from Candida species.FEMS Yeast Res. 2015 Jun;15(4):fov016. doi: 10.1093/femsyr/fov016. Epub 2015 Apr 15. FEMS Yeast Res. 2015. PMID: 25878052 Review.
-
Candida and candidaemia. Susceptibility and epidemiology.Dan Med J. 2013 Nov;60(11):B4698. Dan Med J. 2013. PMID: 24192246 Review.
Cited by
-
Molecular Dissection of Crz1 and Its Dynamic Subcellular Localization in Cryptococcus neoformans.J Fungi (Basel). 2023 Feb 14;9(2):252. doi: 10.3390/jof9020252. J Fungi (Basel). 2023. PMID: 36836365 Free PMC article.
-
Evaluation of Virulence Factors In vitro, Resistance to Osmotic Stress and Antifungal Susceptibility of Candida tropicalis Isolated from the Coastal Environment of Northeast Brazil.Front Microbiol. 2016 Nov 15;7:1783. doi: 10.3389/fmicb.2016.01783. eCollection 2016. Front Microbiol. 2016. PMID: 27895625 Free PMC article.
-
Development of a yeast cell based method for efficient screening of high yield tacrolimus production strain.3 Biotech. 2024 Jan;14(1):26. doi: 10.1007/s13205-023-03870-y. Epub 2023 Dec 31. 3 Biotech. 2024. PMID: 38169568 Free PMC article.
-
An Overlooked and Underrated Endemic Mycosis-Talaromycosis and the Pathogenic Fungus Talaromyces marneffei.Clin Microbiol Rev. 2023 Mar 23;36(1):e0005122. doi: 10.1128/cmr.00051-22. Epub 2023 Jan 17. Clin Microbiol Rev. 2023. PMID: 36648228 Free PMC article. Review.
-
Evaluation of a Novel FKS1 R1354H Mutation Associated with Caspofungin Resistance in Candida auris Using the CRISPR-Cas9 System.J Fungi (Basel). 2023 Apr 29;9(5):529. doi: 10.3390/jof9050529. J Fungi (Basel). 2023. PMID: 37233240 Free PMC article.
References
-
- Arendrup MC, Bruun B, Christensen JJ, Fuursted K, Johansen HK, Kjaeldgaard P, Knudsen JD, Kristensen L, Moller J, Nielsen L, Rosenvinge FS, Roder B, Schonheyder HC, Thomsen MK, Truberg K. 2011. National surveillance of fungemia in Denmark (2004 to 2009). J. Clin. Microbiol. 49:325–334. 10.1128/JCM.01811-10 - DOI - PMC - PubMed
-
- Arendrup MC, Fuursted K, Gahrn-Hansen B, Schonheyder HC, Knudsen JD, Jensen IM, Bruun B, Christensen JJ, Johansen HK. 2008. Semi-national surveillance of fungaemia in Denmark 2004-2006: increasing incidence of fungaemia and numbers of isolates with reduced azole susceptibility. Clin. Microbiol. Infect. 14:487–494. 10.1111/j.1469-0691.2008.01954.x - DOI - PubMed
-
- Andes DR, Safdar N, Baddley JW, Playford G, Reboli AC, Rex JH, Sobel JD, Pappas PG, Kullberg BJ, Mycoses Study Group 2012. Impact of treatment strategy on outcomes in patients with candidemia and other forms of invasive candidiasis: a patient-level quantitative review of randomized trials. Clin. Infect. Dis. 54:1110–1122. 10.1093/cid/cis021 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
