

Origin and pharmacological modulation of tumor-associated regulatory dendritic cells
- PMID: 24443321
- PMCID: PMC4030414
- DOI: 10.1002/ijc.28590
Origin and pharmacological modulation of tumor-associated regulatory dendritic cells
Abstract
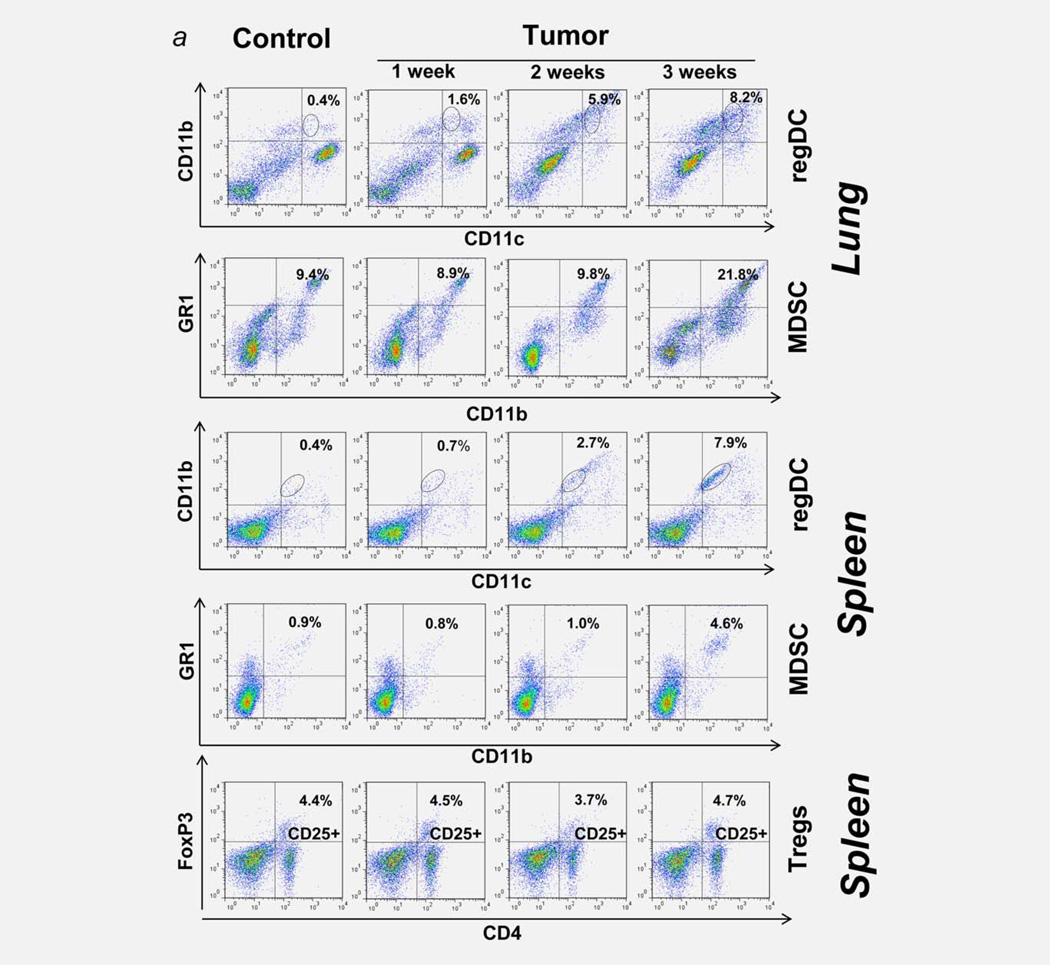
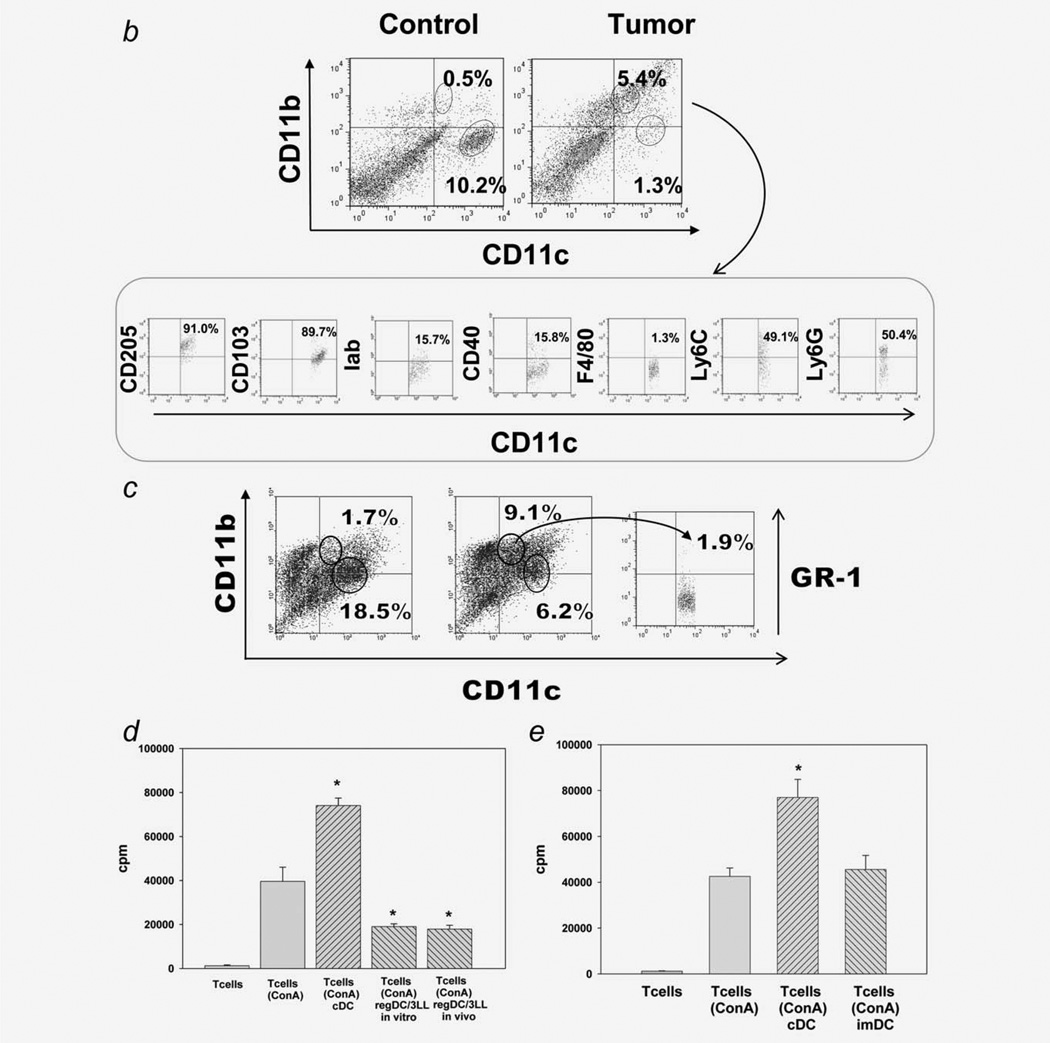
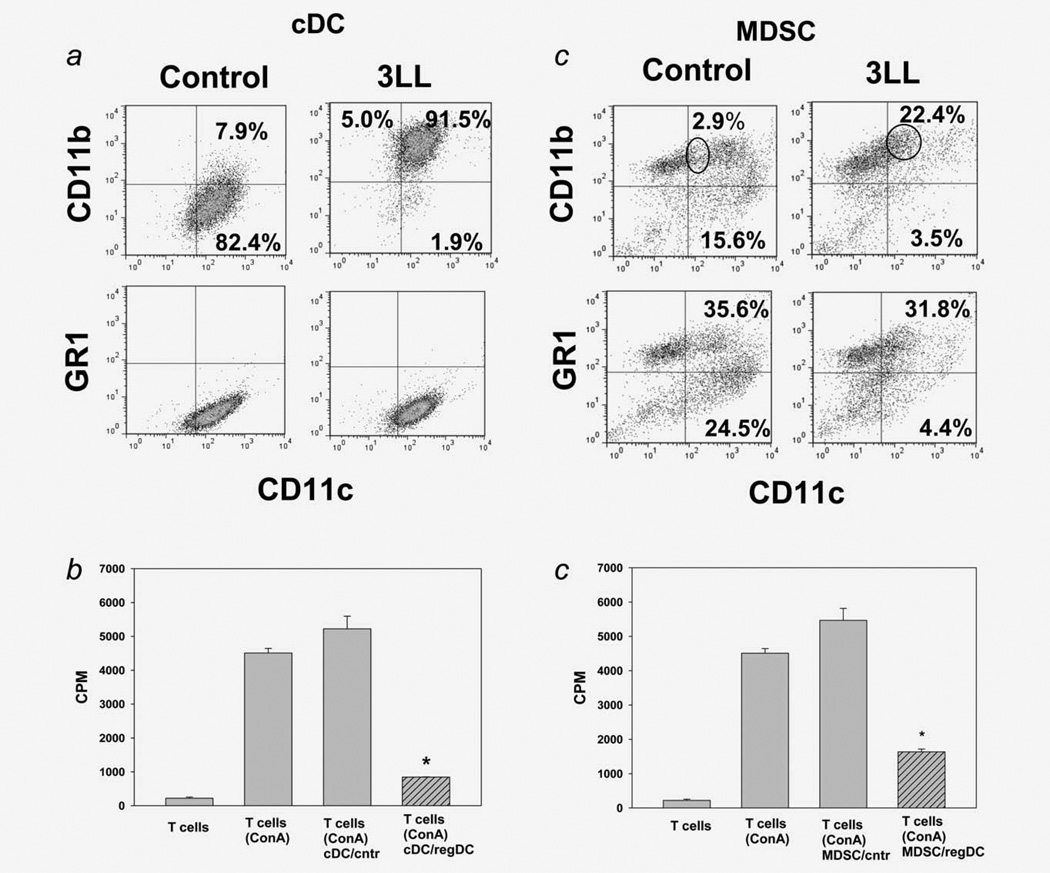
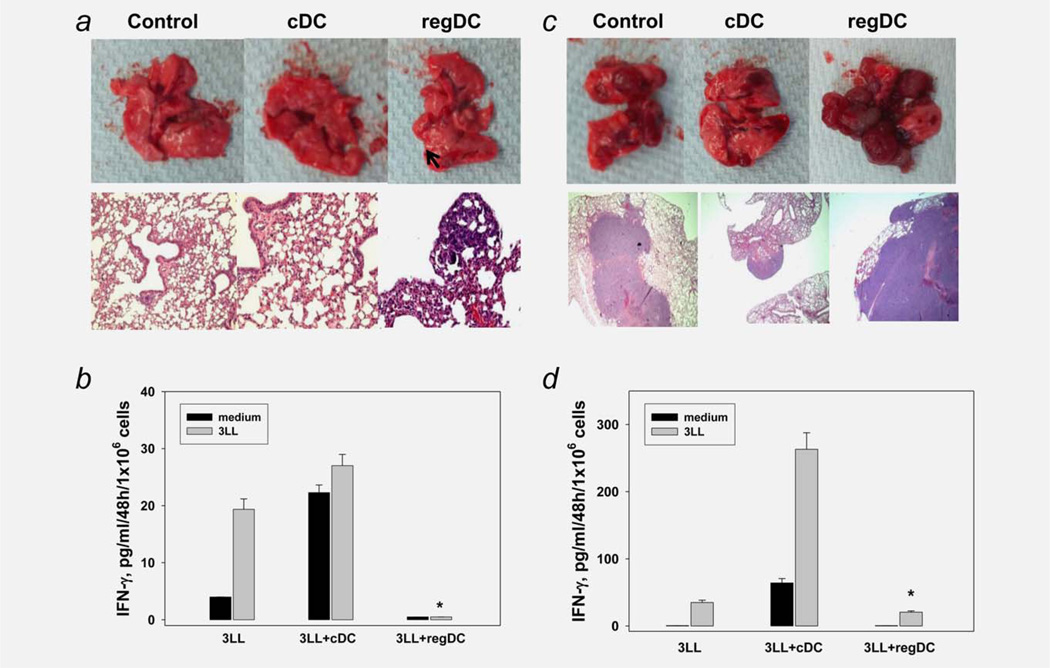
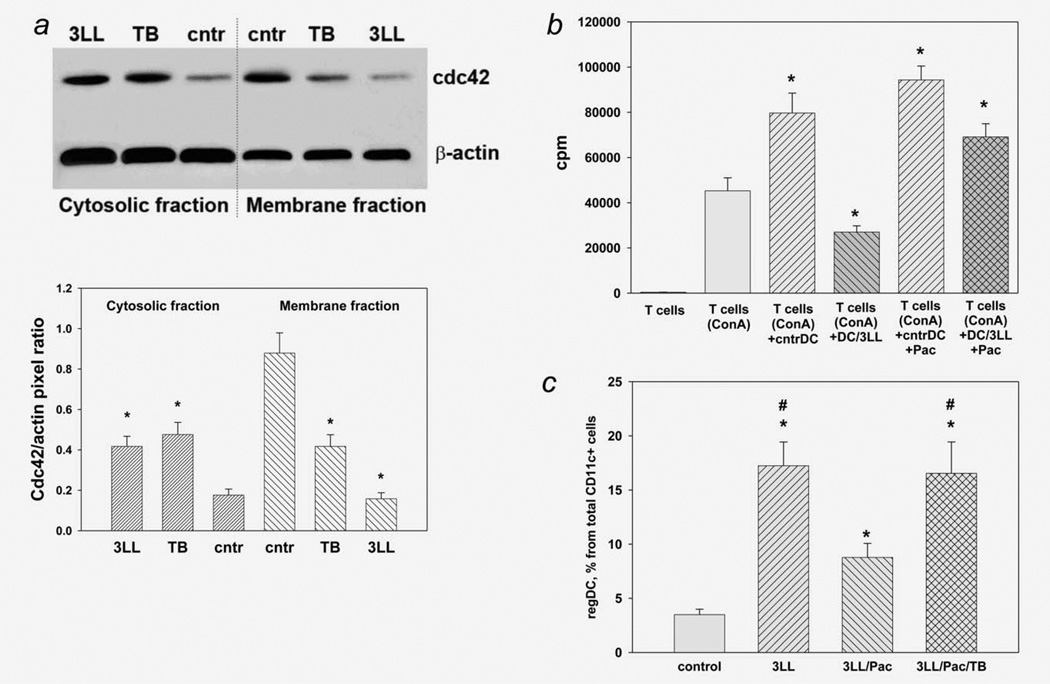
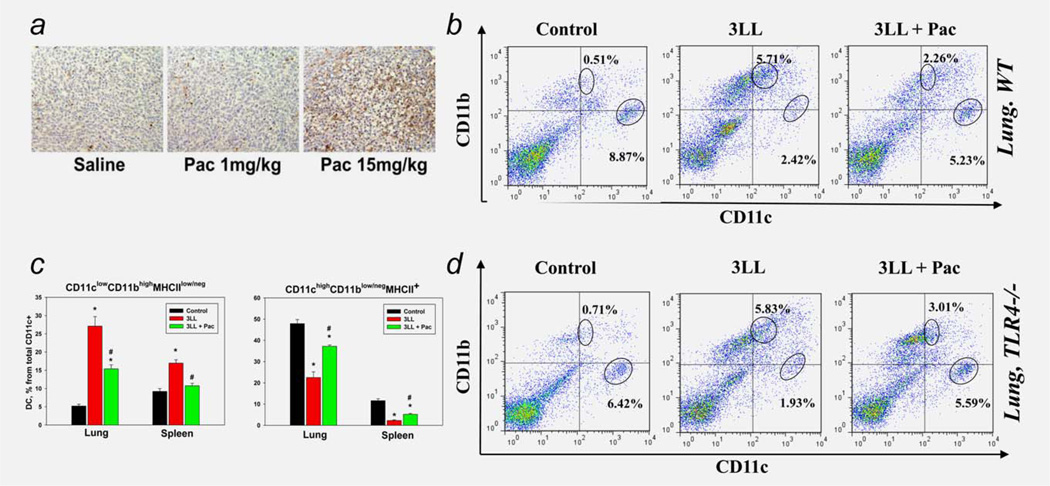
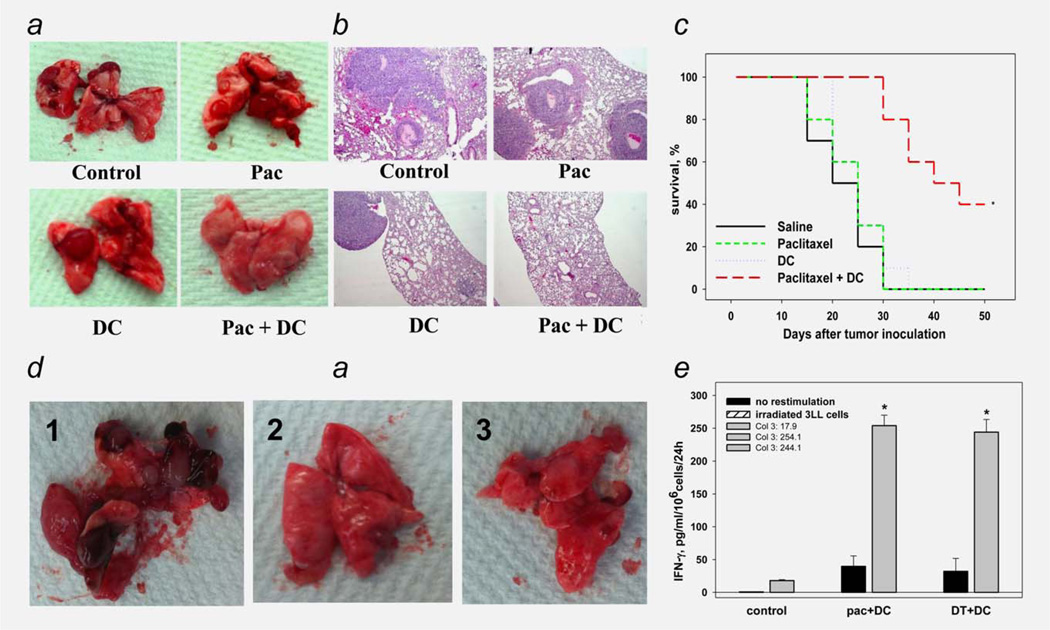
Protumorigenic activity of immune regulatory cells has been proven to play a major role in precluding immunosurveillance and limiting the efficacy of anticancer therapies. Although several approaches have been offered to deplete myeloid-derived suppressor cells (MDSC) and regulatory T cells, there are no data on how to control suppressive dendritic cell (DC) accumulation or function in the tumor environment. Although immunosuppressive function of DC in cancer was implicated to immature and plasmacytoid DC, details of how conventional DC (cDC) develop immunosuppressive properties remain less understood. Here, we show that the development of lung cancer in mice was associated with fast accumulation of regulatory DC (regDC) prior to the appearance of MDSC. Using the in vitro and in vivo approaches, we demonstrated that (i)both cDC and MDSC could be polarized into protumor regDC in the lung cancer environment; (ii) cDC → regDC polarization was mediated by the small Rho GTPase signaling, which could be controlled by noncytotoxic doses of paclitaxel; and (iii) prevention of regDC appearance increased the antitumor potential of DC vaccine in lung cancer. These findings not only bring new players to the family of myeloid regulatory cells and provide new targets for cancer therapy, but offer novel insights into the immunomodulatory capacity of chemotherapeutic agents used in low, noncytotoxic doses.
Keywords: MDSC; dendritic cells; small Rho GTPases; taxol; tumor microenvironment.
© 2013 UICC.
Figures
Similar articles
-
The inflammasome component NLRP3 impairs antitumor vaccine by enhancing the accumulation of tumor-associated myeloid-derived suppressor cells.Cancer Res. 2010 Dec 15;70(24):10161-9. doi: 10.1158/0008-5472.CAN-10-1921. Cancer Res. 2010. PMID: 21159638 Free PMC article.
-
Enhancement of antitumor immunity in lung cancer by targeting myeloid-derived suppressor cell pathways.Cancer Res. 2013 Nov 15;73(22):6609-20. doi: 10.1158/0008-5472.CAN-13-0987. Epub 2013 Oct 1. Cancer Res. 2013. PMID: 24085788 Free PMC article.
-
Antitumor effect of paclitaxel is mediated by inhibition of myeloid-derived suppressor cells and chronic inflammation in the spontaneous melanoma model.J Immunol. 2013 Mar 1;190(5):2464-71. doi: 10.4049/jimmunol.1202781. Epub 2013 Jan 28. J Immunol. 2013. PMID: 23359505 Free PMC article.
-
Adjuvants and myeloid-derived suppressor cells: enemies or allies in therapeutic cancer vaccination.Hum Vaccin Immunother. 2014;10(11):3251-60. doi: 10.4161/hv.29847. Hum Vaccin Immunother. 2014. PMID: 25483674 Free PMC article. Review.
-
Breaking immunotolerance of tumors: a new perspective for dendritic cell therapy.J Immunotoxicol. 2014 Oct;11(4):311-8. doi: 10.3109/1547691X.2013.865094. Epub 2014 Feb 4. J Immunotoxicol. 2014. PMID: 24495309 Review.
Cited by
-
Targeting myeloid regulators by paclitaxel-loaded enzymatically degradable nanocups.Nanoscale. 2018 Sep 27;10(37):17990-18000. doi: 10.1039/c8nr04437f. Nanoscale. 2018. PMID: 30226240 Free PMC article.
-
The role of tumor microenvironment on cancer stem cell fate in solid tumors.Cell Commun Signal. 2023 Jun 16;21(1):143. doi: 10.1186/s12964-023-01129-w. Cell Commun Signal. 2023. PMID: 37328876 Free PMC article. Review.
-
Stimulation of regulatory dendritic cells suppresses cytotoxic T cell function and alleviates DEN-induced liver injury, fibrosis and hepatocellular carcinoma.Front Immunol. 2025 Apr 8;16:1565486. doi: 10.3389/fimmu.2025.1565486. eCollection 2025. Front Immunol. 2025. PMID: 40264769 Free PMC article.
-
Nano-gold corking and enzymatic uncorking of carbon nanotube cups.J Am Chem Soc. 2015 Jan 21;137(2):675-84. doi: 10.1021/ja511843w. Epub 2015 Jan 7. J Am Chem Soc. 2015. PMID: 25530234 Free PMC article.
-
Dendritic Cells in Shaping Anti-Tumor T Cell Response.Cancers (Basel). 2024 Jun 13;16(12):2211. doi: 10.3390/cancers16122211. Cancers (Basel). 2024. PMID: 38927916 Free PMC article. Review.
References
-
- Spano D, Zollo M. Tumor microenvironment: a main actor in the metastasis process. Clin Exp Metastasis. 2012;29:381–395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
