

Cell-selective knockout and 3D confocal image analysis reveals separate roles for astrocyte-and endothelial-derived CCL2 in neuroinflammation
- PMID: 24444311
- PMCID: PMC3906899
- DOI: 10.1186/1742-2094-11-10
Cell-selective knockout and 3D confocal image analysis reveals separate roles for astrocyte-and endothelial-derived CCL2 in neuroinflammation
Abstract
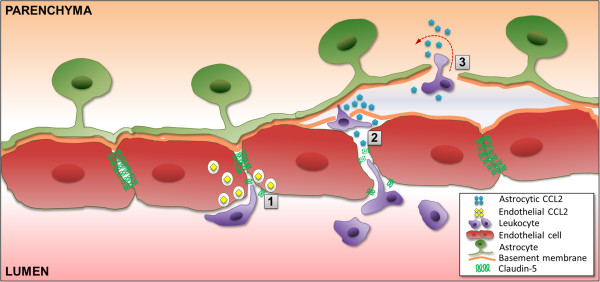
Background: Expression of chemokine CCL2 in the normal central nervous system (CNS) is nearly undetectable, but is significantly upregulated and drives neuroinflammation during experimental autoimmune encephalomyelitis (EAE), an animal model of multiple sclerosis which is considered a contributing factor in the human disease. As astrocytes and brain microvascular endothelial cells (BMEC) forming the blood-brain barrier (BBB) are sources of CCL2 in EAE and other neuroinflammatory conditions, it is unclear if one or both CCL2 pools are critical to disease and by what mechanism(s).
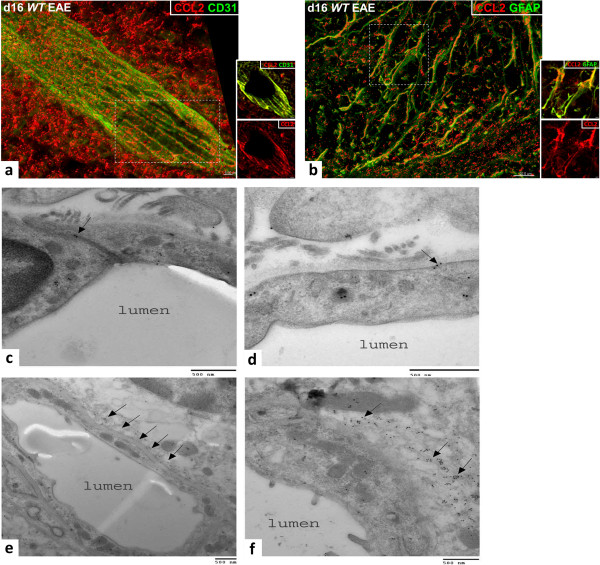
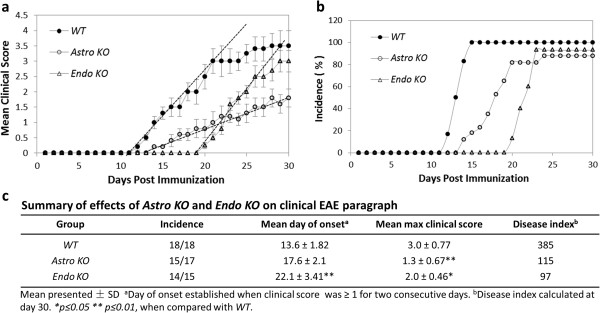
Methods: Mice with selective CCL2 gene knockout (KO) in astrocytes (Astro KO) or endothelial cells (Endo KO) were used to evaluate the respective contributions of these sources to neuroinflammation, i.e., clinical disease progression, BBB damage, and parenchymal leukocyte invasion in a myelin oligodendrocyte glycoprotein peptide (MOG35-55)-induced EAE model. High-resolution 3-dimensional (3D) immunofluorescence confocal microscopy and colloidal gold immuno-electron microscopy were employed to confirm sites of CCL2 expression, and 3D immunofluorescence confocal microscopy utilized to assess inflammatory responses along the CNS microvasculature.
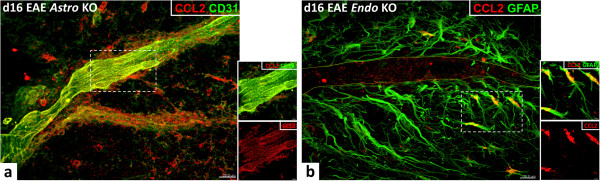
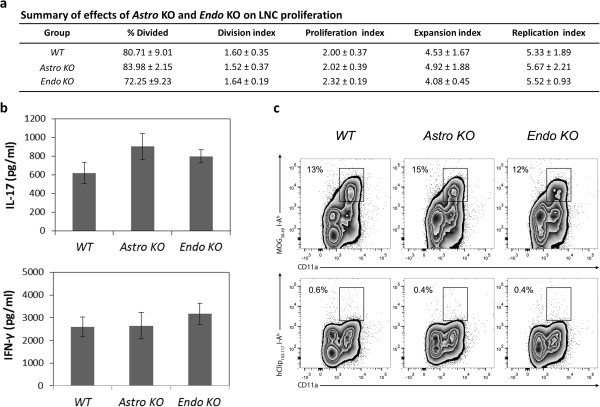
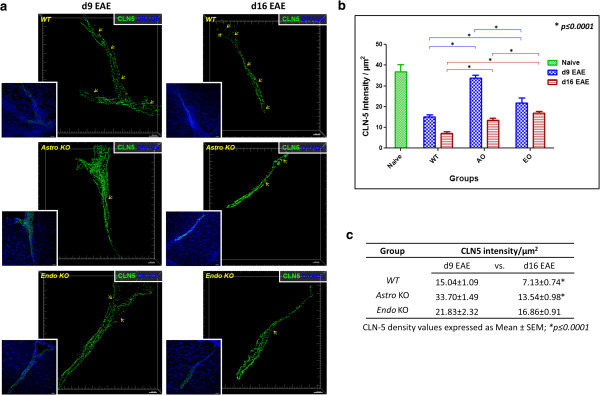
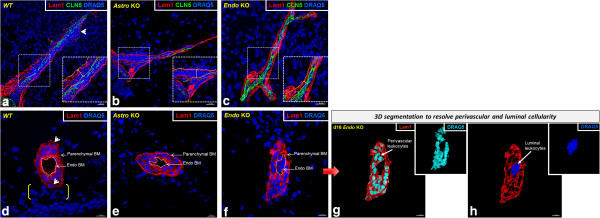
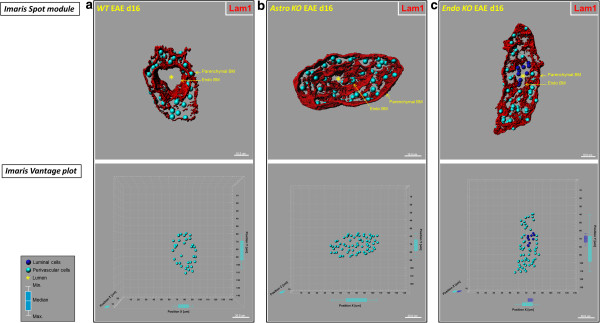
Results: Cell-selective loss of CCL2 immunoreactivity was demonstrated in the respective KO mice. Compared to wild-type (WT) mice, Astro KO mice showed reduced EAE severity but similar onset, while Endo KO mice displayed near normal severity but significantly delayed onset. Neither of the KO mice showed deficits in T cell proliferation, or IL-17 and IFN-γ production, following MOG35-55 exposure in vitro, or altered MOG-major histocompatibility complex class II tetramer binding. 3D confocal imaging further revealed distinct actions of the two CCL2 pools in the CNS. Astro KOs lacked the CNS leukocyte penetration and disrupted immunostaining of CLN-5 at the BBB seen during early EAE in WT mice, while Endo KOs uniquely displayed leukocytes stalled in the microvascular lumen.
Conclusions: These results point to astrocyte and endothelial pools of CCL2 each regulating different stages of neuroinflammation in EAE, and carry implications for drug delivery in neuroinflammatory disease.
Figures
Similar articles
-
Appearance of claudin-5+ leukocytes in the central nervous system during neuroinflammation: a novel role for endothelial-derived extracellular vesicles.J Neuroinflammation. 2016 Nov 16;13(1):292. doi: 10.1186/s12974-016-0755-8. J Neuroinflammation. 2016. PMID: 27852330 Free PMC article.
-
Kinin B2 receptor regulates chemokines CCL2 and CCL5 expression and modulates leukocyte recruitment and pathology in experimental autoimmune encephalomyelitis (EAE) in mice.J Neuroinflammation. 2008 Nov 5;5:49. doi: 10.1186/1742-2094-5-49. J Neuroinflammation. 2008. PMID: 18986535 Free PMC article.
-
Rescue from acute neuroinflammation by pharmacological chemokine-mediated deviation of leukocytes.J Neuroinflammation. 2012 Oct 25;9:243. doi: 10.1186/1742-2094-9-243. J Neuroinflammation. 2012. PMID: 23095573 Free PMC article.
-
Regulation of autoimmune-mediated neuroinflammation by endothelial cells.Eur J Immunol. 2024 Apr;54(4):e2350482. doi: 10.1002/eji.202350482. Epub 2024 Feb 9. Eur J Immunol. 2024. PMID: 38335316 Review.
-
The Rise of Pluripotent Stem Cell-Derived Glia Models of Neuroinflammation.Neurol Int. 2025 Jan 13;17(1):6. doi: 10.3390/neurolint17010006. Neurol Int. 2025. PMID: 39852770 Free PMC article. Review.
Cited by
-
Cardiac glycosides target barrier inflammation of the vasculature, meninges and choroid plexus.Commun Biol. 2021 Feb 26;4(1):260. doi: 10.1038/s42003-021-01787-x. Commun Biol. 2021. PMID: 33637884 Free PMC article.
-
Control of autoimmune CNS inflammation by astrocytes.Semin Immunopathol. 2015 Nov;37(6):625-38. doi: 10.1007/s00281-015-0515-3. Epub 2015 Jul 30. Semin Immunopathol. 2015. PMID: 26223505 Free PMC article. Review.
-
Aberrant Cerebral Iron Trafficking Co-morbid With Chronic Inflammation: Molecular Mechanisms and Pharmacologic Intervention.Front Neurol. 2022 Mar 15;13:855751. doi: 10.3389/fneur.2022.855751. eCollection 2022. Front Neurol. 2022. PMID: 35370907 Free PMC article. Review.
-
Astrogliosis in multiple sclerosis and neuro-inflammation: what role for the notch pathway?Front Immunol. 2023 Oct 23;14:1254586. doi: 10.3389/fimmu.2023.1254586. eCollection 2023. Front Immunol. 2023. PMID: 37936690 Free PMC article. Review.
-
Mechanisms of astrocyte aging in reactivity and disease.Mol Neurodegener. 2025 Feb 21;20(1):21. doi: 10.1186/s13024-025-00810-7. Mol Neurodegener. 2025. PMID: 39979986 Free PMC article. Review.
References
-
- Mantovani A, Sozzani S, Bottazzi B, Peri G, Sciacca FL, Locati M, Colotta F. Monocyte chemotactic protein-1 (MCP-1): signal transduction and involvement in the regulation of macrophage traffic in normal and neoplastic tissues. Adv Exp Med Biol. 1993;351:47–54. doi: 10.1007/978-1-4615-2952-1_6. - DOI - PubMed
-
- Bennett JL, Elhofy A, Canto MC, Tani M, Ransohoff RM, Karpus WJ. CCL2 transgene expression in the central nervous system directs diffuse infiltration of CD45(high)CD11b(+) monocytes and enhanced Theiler’s murine encephalomyelitis virus-induced demyelinating disease. J Neurovirol. 2003;9:623–636. - PMC - PubMed
-
- Toft-Hansen H, Buist R, Sun XJ, Schellenberg A, Peeling J, Owens T. Metalloproteinases control brain inflammation induced by pertussis toxin in mice overexpressing the chemokine CCL2 in the central nervous system. J Immunol. 2006;177:7242–7249. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
