

A reduction in Ptprq associated with specific features of the deafness phenotype of the miR-96 mutant mouse diminuendo
- PMID: 24446963
- PMCID: PMC4065360
- DOI: 10.1111/ejn.12484
A reduction in Ptprq associated with specific features of the deafness phenotype of the miR-96 mutant mouse diminuendo
Abstract
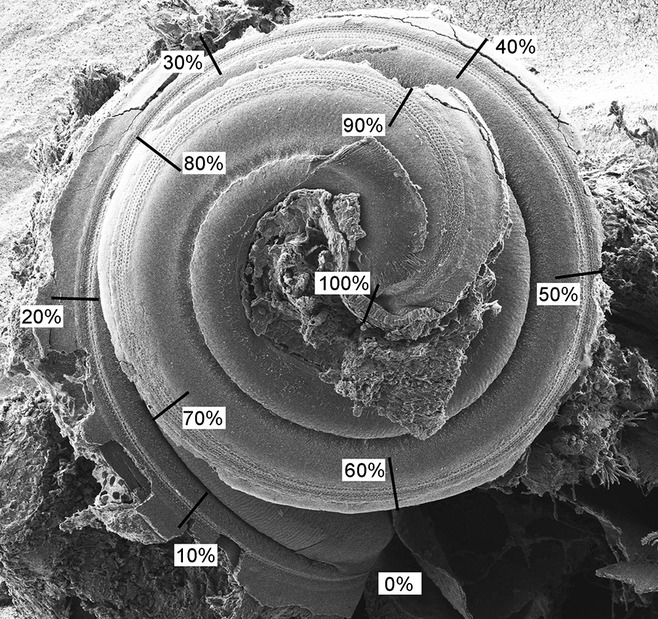
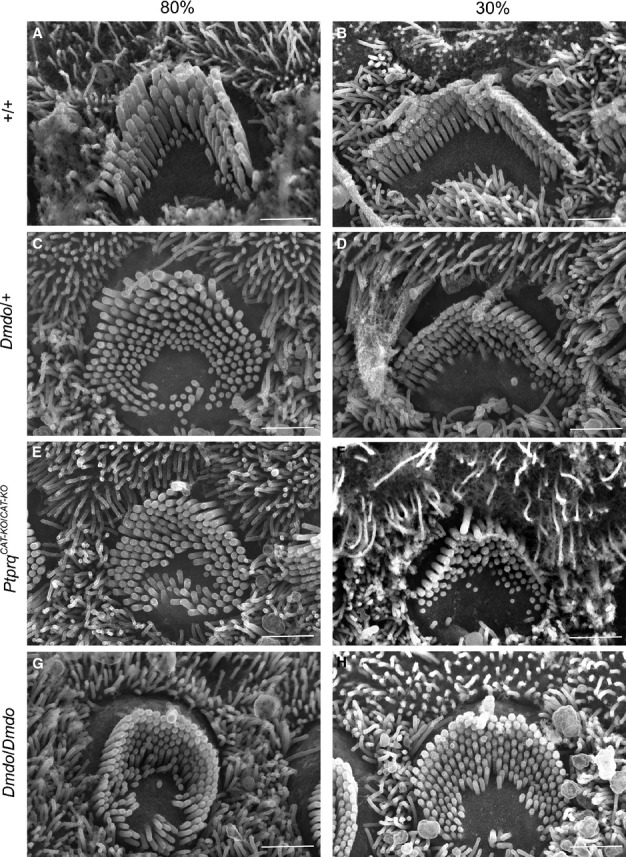
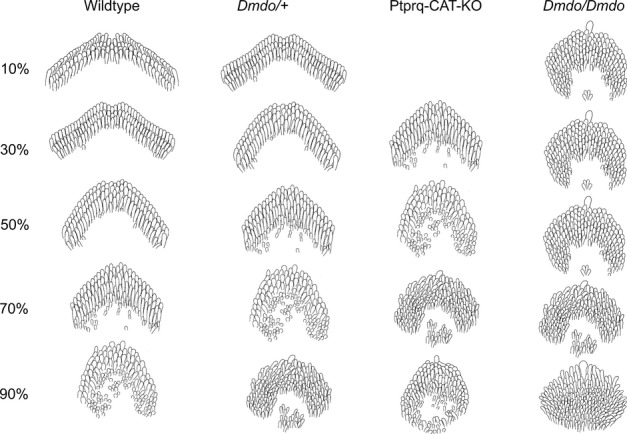
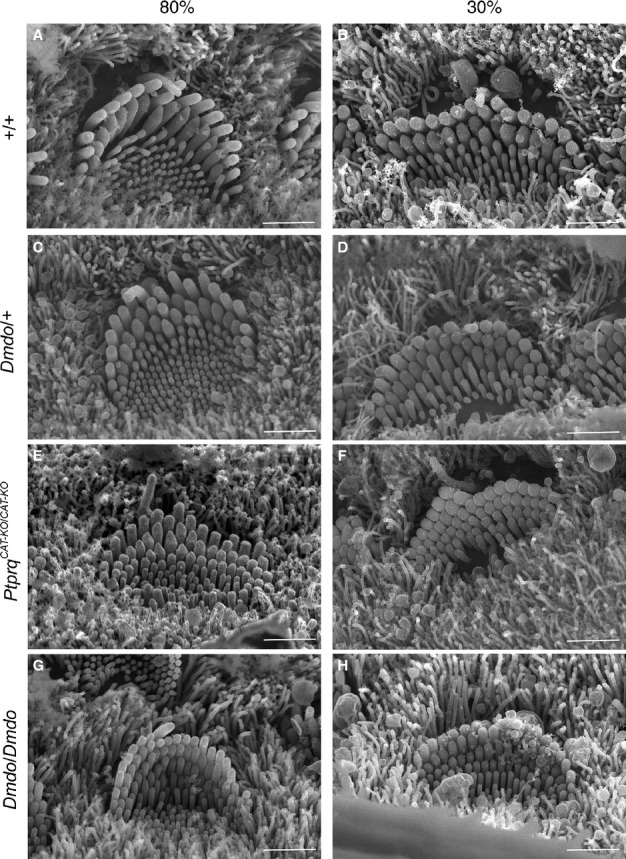
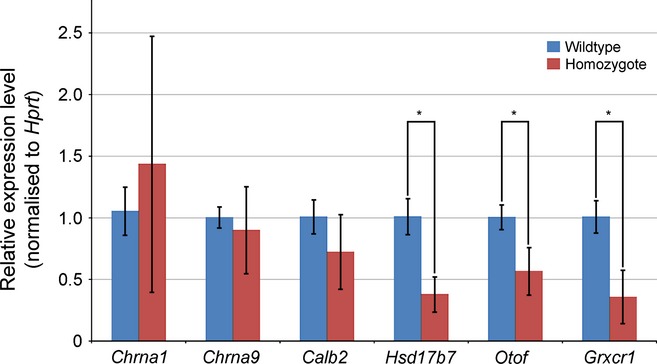
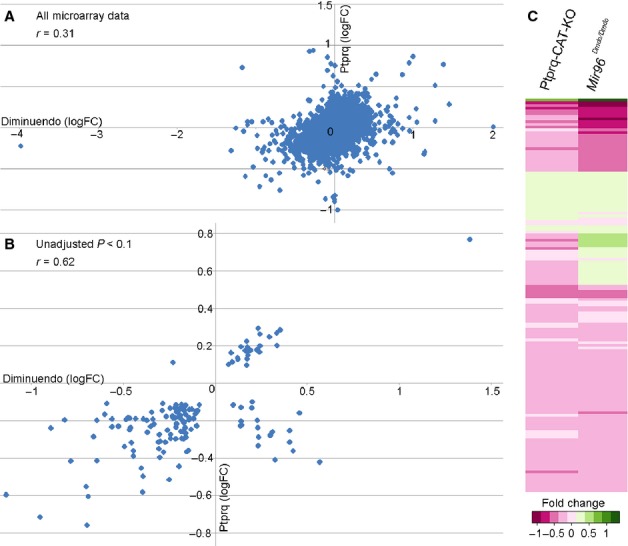
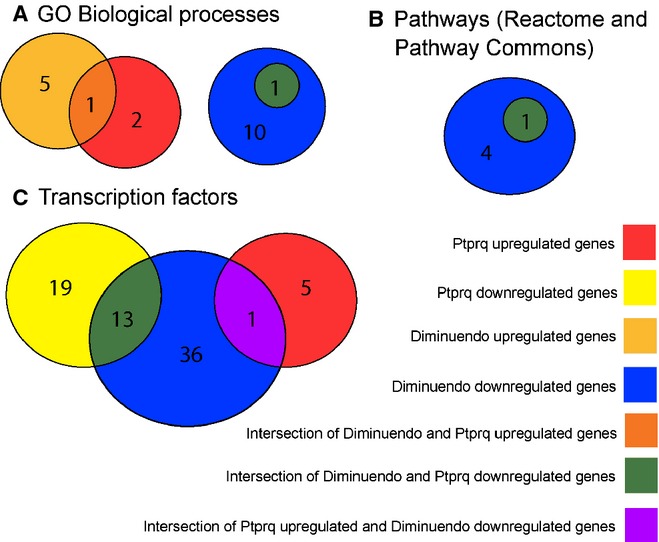
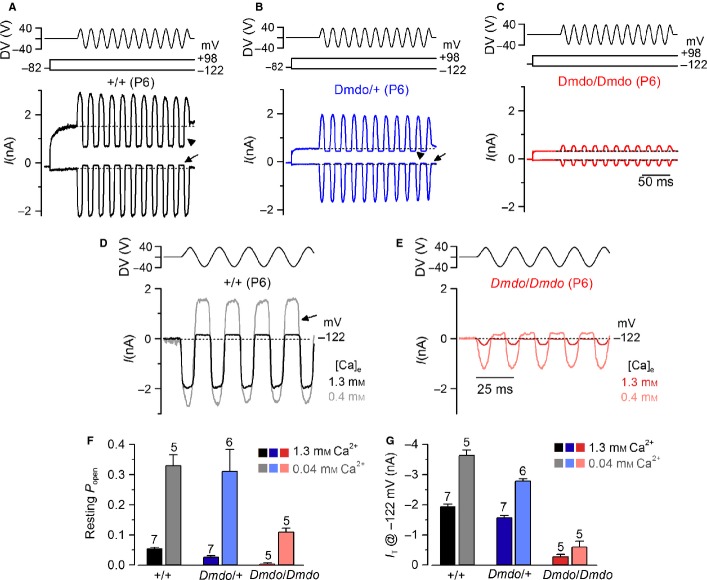
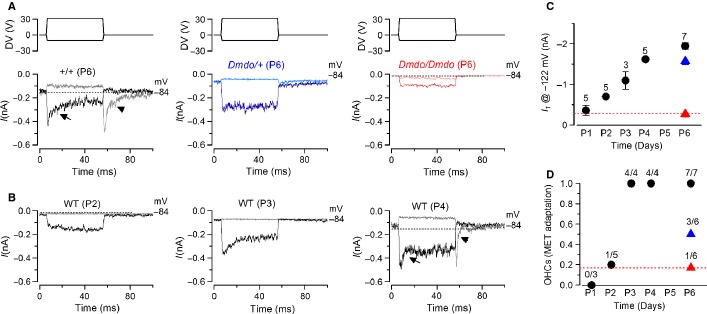
miR-96 is a microRNA, a non-coding RNA gene which regulates a wide array of downstream genes. The miR-96 mouse mutant diminuendo exhibits deafness and arrested hair cell functional and morphological differentiation. We have previously shown that several genes are markedly downregulated in the diminuendo organ of Corti; one of these is Ptprq, a gene known to be important for maturation and maintenance of hair cells. In order to study the contribution that downregulation of Ptprq makes to the diminuendo phenotype, we carried out microarrays, scanning electron microscopy and single hair cell electrophysiology to compare diminuendo mutants (heterozygous and homozygous) with mice homozygous for a functional null allele of Ptprq. In terms of both morphology and electrophysiology, the auditory phenotype of mice lacking Ptprq resembles that of diminuendo heterozygotes, while diminuendo homozygotes are more severely affected. A comparison of transcriptomes indicates there is a broad similarity between diminuendo homozygotes and Ptprq-null mice. The reduction in Ptprq observed in diminuendo mice appears to be a major contributor to the morphological, transcriptional and electrophysiological phenotype, but does not account for the complete diminuendo phenotype.
Keywords: ear development; hereditary hearing loss; knockout and transgenic m; molecular genetics; sensory hair cells.
© 2014 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures


References
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B. 1995;57:289–300.
-
- Bolstad B. 2001. Probe level quantile normalization of high density oligonucleotide array data. Available at http://bmbolstad.com/stuff/qnorm.pdf.
-
- Bolstad BM, Irizarry RA, Astrand M, Speed TP. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics. 2003;19:185–193. - PubMed
-
- Bosher SK, Warren RL. Very low calcium content of cochlear endolymph, an extracellular fluid. Nature. 1978;273:377–378. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
