

Mechanical motion promotes expression of Prg4 in articular cartilage via multiple CREB-dependent, fluid flow shear stress-induced signaling pathways
- PMID: 24449269
- PMCID: PMC3909787
- DOI: 10.1101/gad.231969.113
Mechanical motion promotes expression of Prg4 in articular cartilage via multiple CREB-dependent, fluid flow shear stress-induced signaling pathways
Abstract
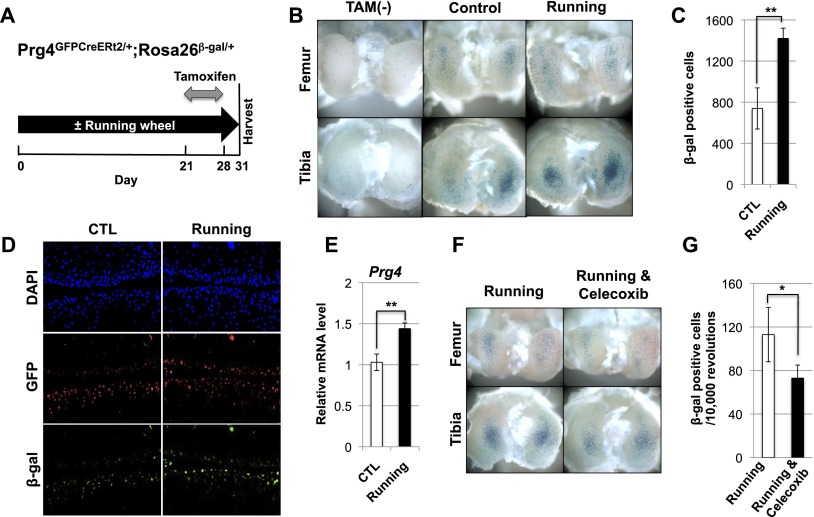
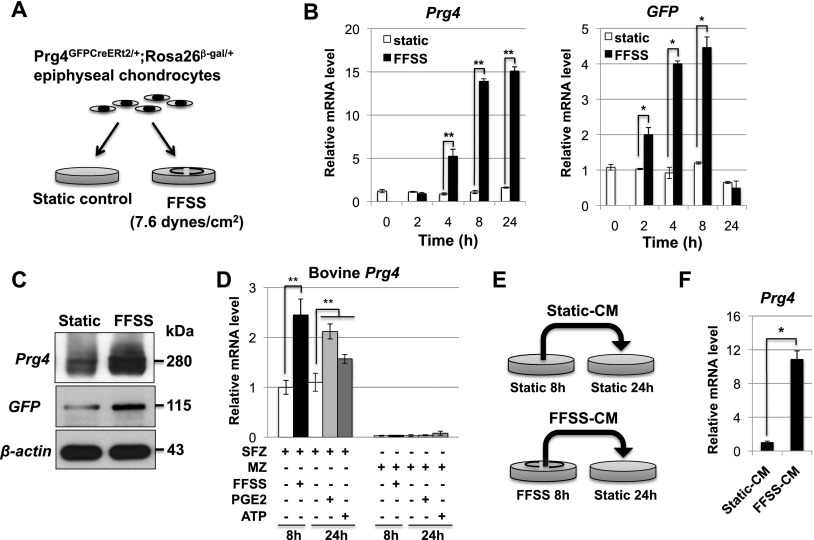
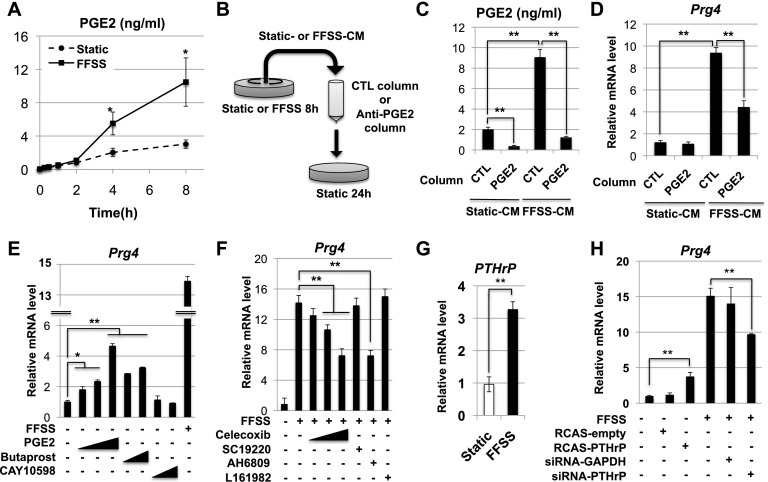
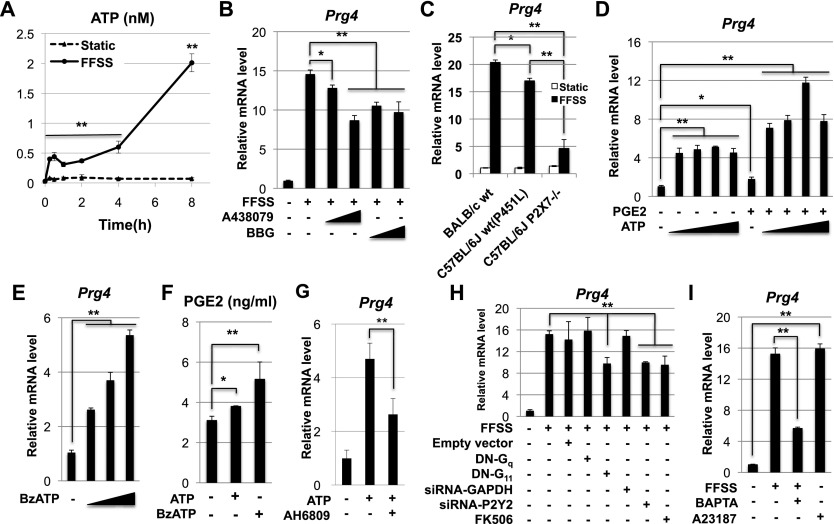
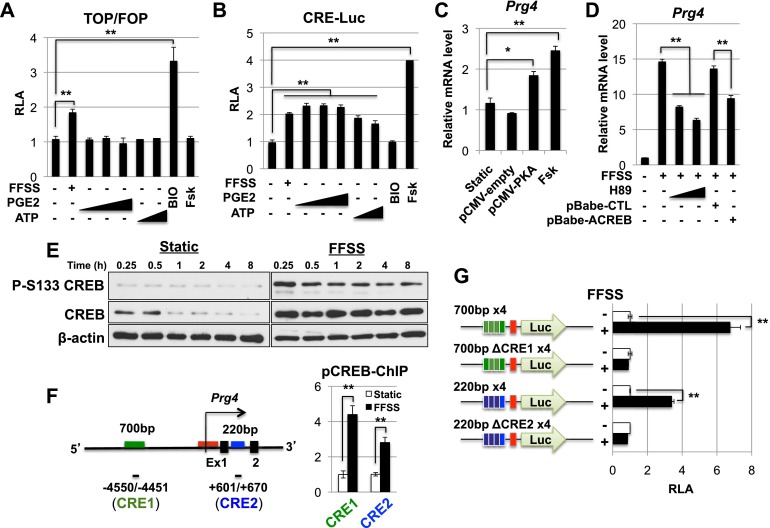
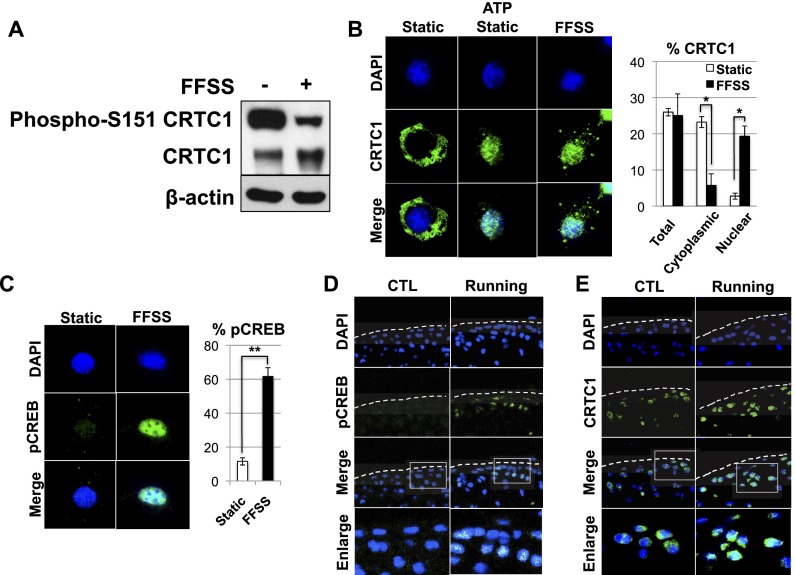
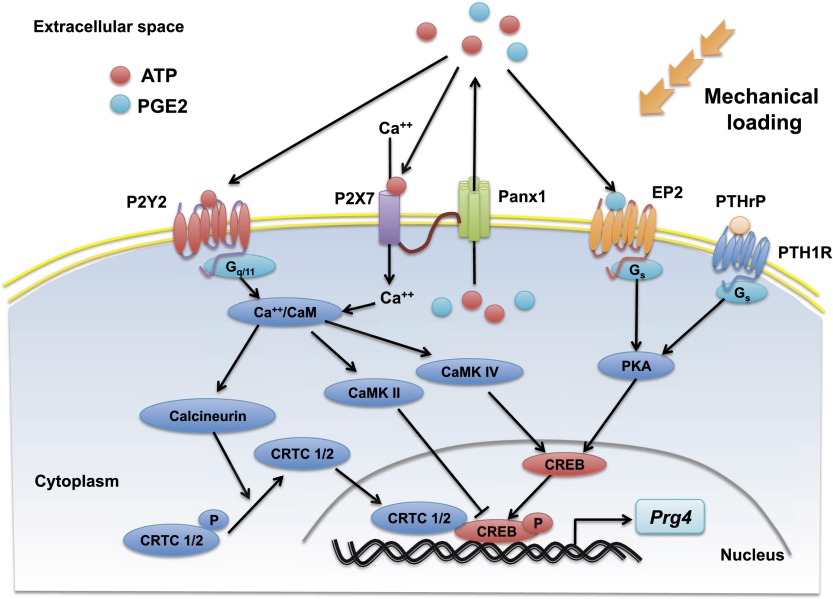
Lubricin is a secreted proteoglycan encoded by the Prg4 locus that is abundantly expressed by superficial zone articular chondrocytes and has been noted to both be sensitive to mechanical loading and protect against the development of osteoarthritis. In this study, we document that running induces maximal expression of Prg4 in the superficial zone of knee joint articular cartilage in a COX-2-dependent fashion, which correlates with augmented levels of phospho-S133 CREB and increased nuclear localization of CREB-regulated transcriptional coactivators (CRTCs) in this tissue. Furthermore, we found that fluid flow shear stress (FFSS) increases secretion of extracellular PGE2, PTHrP, and ATP (by epiphyseal chondrocytes), which together engage both PKA- and Ca(++)-regulated signaling pathways that work in combination to promote CREB-dependent induction of Prg4, specifically in superficial zone articular chondrocytes. Because running and FFSS both boost Prg4 expression in a COX-2-dependent fashion, our results suggest that mechanical motion may induce Prg4 expression in the superficial zone of articular cartilage by engaging the same signaling pathways activated in vitro by FFSS that promote CREB-dependent gene expression in this tissue.
Keywords: CREB; PGE2; PTHrP; Prg4/lubricin; articular cartilage; extracellular ATP.
Figures
Similar articles
-
Wnt/β-catenin signaling contributes to articular cartilage homeostasis through lubricin induction in the superficial zone.Arthritis Res Ther. 2019 Nov 27;21(1):247. doi: 10.1186/s13075-019-2041-5. Arthritis Res Ther. 2019. PMID: 31771658 Free PMC article.
-
Creb5 establishes the competence for Prg4 expression in articular cartilage.Commun Biol. 2021 Mar 12;4(1):332. doi: 10.1038/s42003-021-01857-0. Commun Biol. 2021. PMID: 33712729 Free PMC article.
-
Cartilage-specific deletion of Alk5 gene results in a progressive osteoarthritis-like phenotype in mice.Osteoarthritis Cartilage. 2017 Nov;25(11):1868-1879. doi: 10.1016/j.joca.2017.07.010. Epub 2017 Jul 14. Osteoarthritis Cartilage. 2017. PMID: 28716756 Free PMC article.
-
Regulatory Mechanisms of Prg4 and Gdf5 Expression in Articular Cartilage and Functions in Osteoarthritis.Int J Mol Sci. 2022 Apr 23;23(9):4672. doi: 10.3390/ijms23094672. Int J Mol Sci. 2022. PMID: 35563063 Free PMC article. Review.
-
The Actin Cytoskeleton as a Regulator of Proteoglycan 4.Cartilage. 2025 Sep;16(3):376-387. doi: 10.1177/19476035231223455. Epub 2024 Jan 6. Cartilage. 2025. PMID: 38183234 Free PMC article. Review.
Cited by
-
Mechanisms of tissue degeneration mediated by periostin in spinal degenerative diseases and their implications for pathology and diagnosis: a review.Front Med (Lausanne). 2023 Oct 31;10:1276900. doi: 10.3389/fmed.2023.1276900. eCollection 2023. Front Med (Lausanne). 2023. PMID: 38020106 Free PMC article. Review.
-
Regeneration of joint surface defects by transplantation of allogeneic cartilage: application of iPS cell-derived cartilage and immunogenicity.Inflamm Regen. 2023 Nov 14;43(1):56. doi: 10.1186/s41232-023-00307-0. Inflamm Regen. 2023. PMID: 37964383 Free PMC article. Review.
-
Identification of a Prg4-expressing articular cartilage progenitor cell population in mice.Arthritis Rheumatol. 2015 May;67(5):1261-73. doi: 10.1002/art.39030. Arthritis Rheumatol. 2015. PMID: 25603997 Free PMC article.
-
Biofabrication of in situ Self Assembled 3D Cell Cultures in a Weightlessness Environment Generated using Magnetic Levitation.Sci Rep. 2018 May 8;8(1):7239. doi: 10.1038/s41598-018-25718-9. Sci Rep. 2018. PMID: 29740095 Free PMC article.
-
Vaspin antagonizes high fat-induced bone loss in rats and promotes osteoblastic differentiation in primary rat osteoblasts through Smad-Runx2 signaling pathway.Nutr Metab (Lond). 2020 Jan 22;17:9. doi: 10.1186/s12986-020-0429-5. eCollection 2020. Nutr Metab (Lond). 2020. PMID: 31993071 Free PMC article.
References
-
- Adriouch S, Dox C, Welge V, Seman M, Koch-Nolte F, Haag F 2002. Cutting edge: A natural P451L mutation in the cytoplasmic domain impairs the function of the mouse P2X7 receptor. J Immunol 169: 4108–4112 - PubMed
-
- Bahabri SA, Suwairi WM, Laxer RM, Polinkovsky A, Dalaan AA, Warman ML 1998. The camptodactyly-arthropathy-coxa vara-pericarditis syndrome: Clinical features and genetic mapping to human chromosome 1. Arthritis Rheum 41: 730–735 - PubMed
-
- Bao L, Locovei S, Dahl G 2004. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett 572: 65–68 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous