

Epigenetic reprogramming modulates malignant properties of human liver cancer
- PMID: 24449497
- PMCID: PMC4043911
- DOI: 10.1002/hep.27026
Epigenetic reprogramming modulates malignant properties of human liver cancer
Abstract
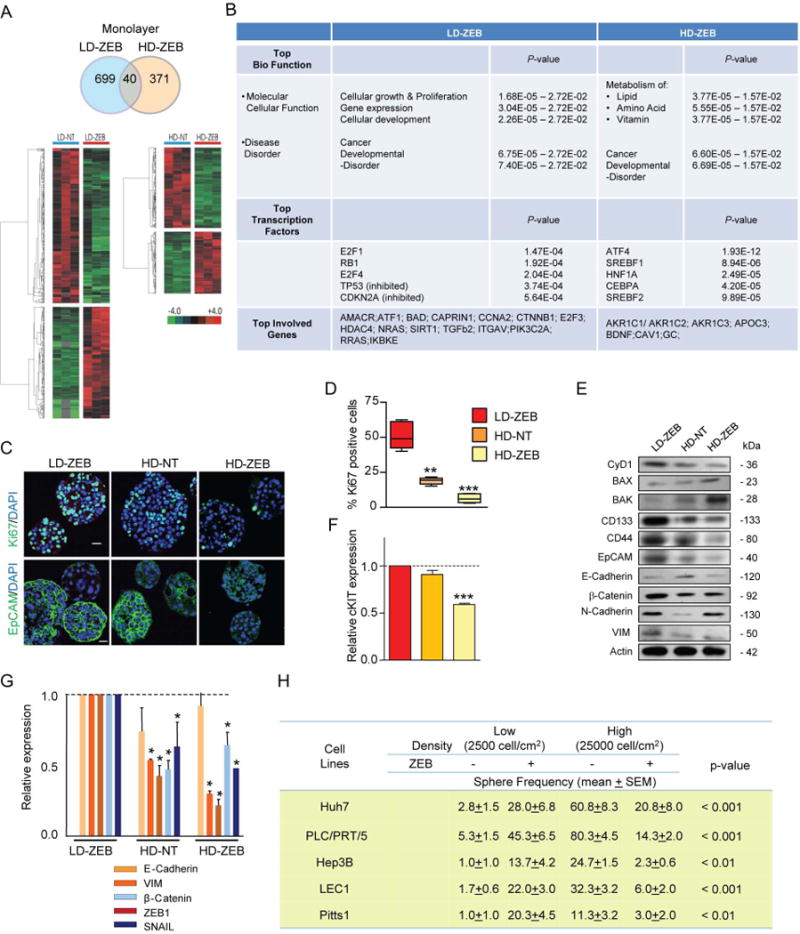
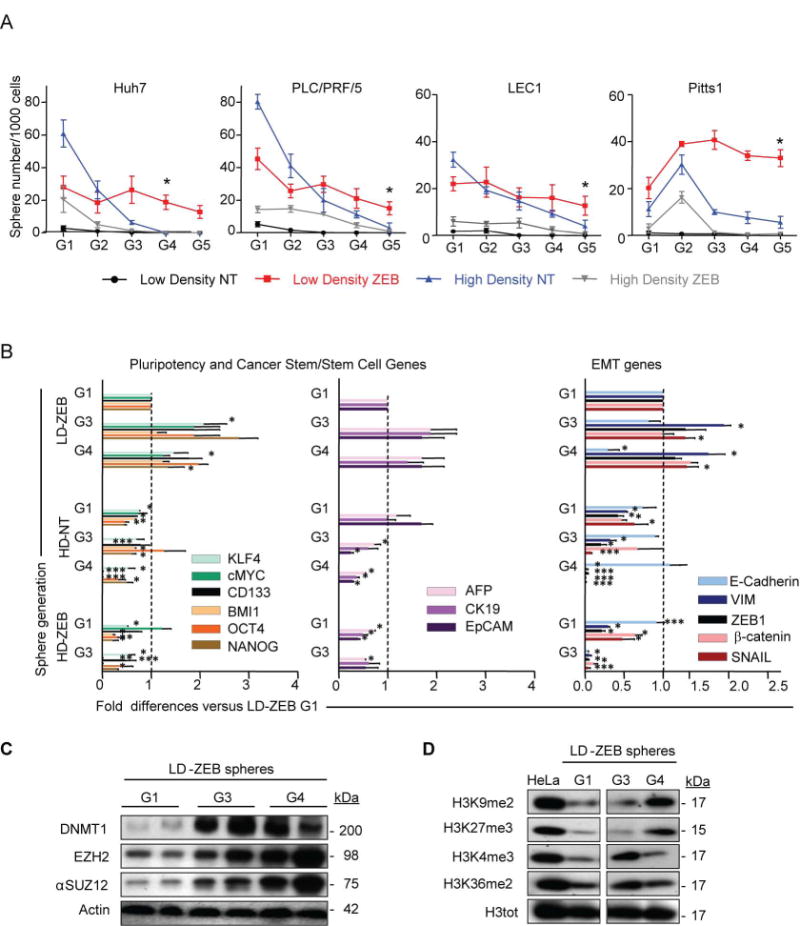
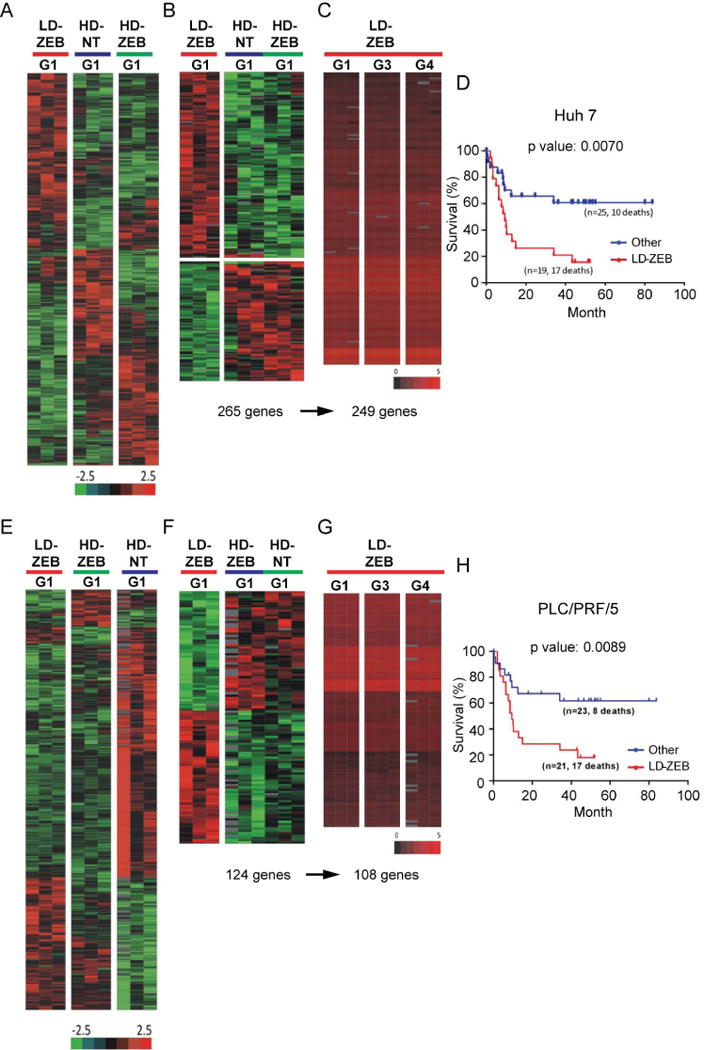
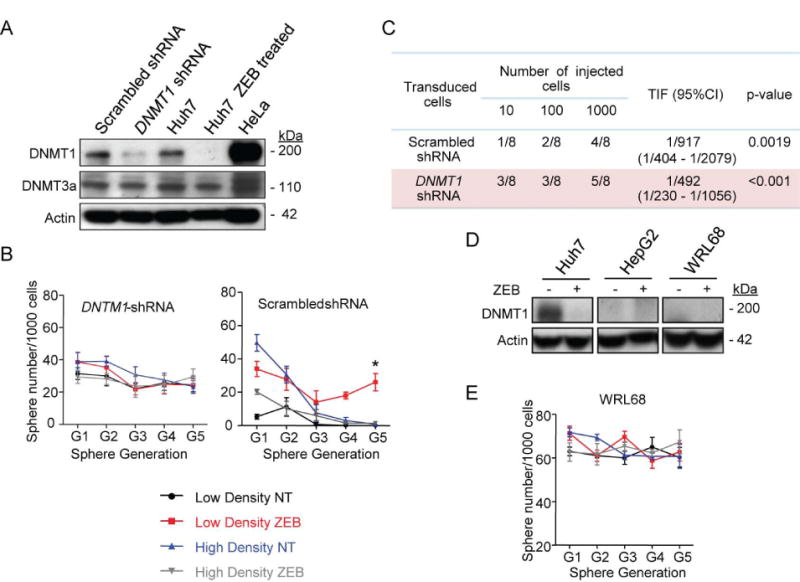
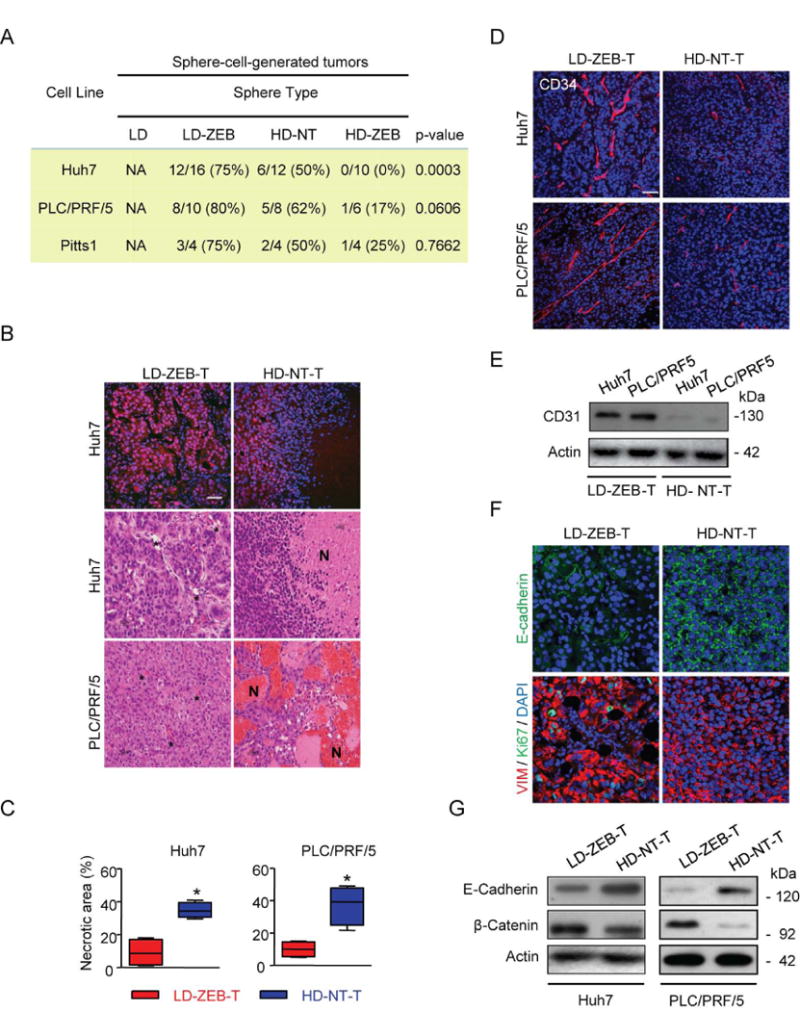
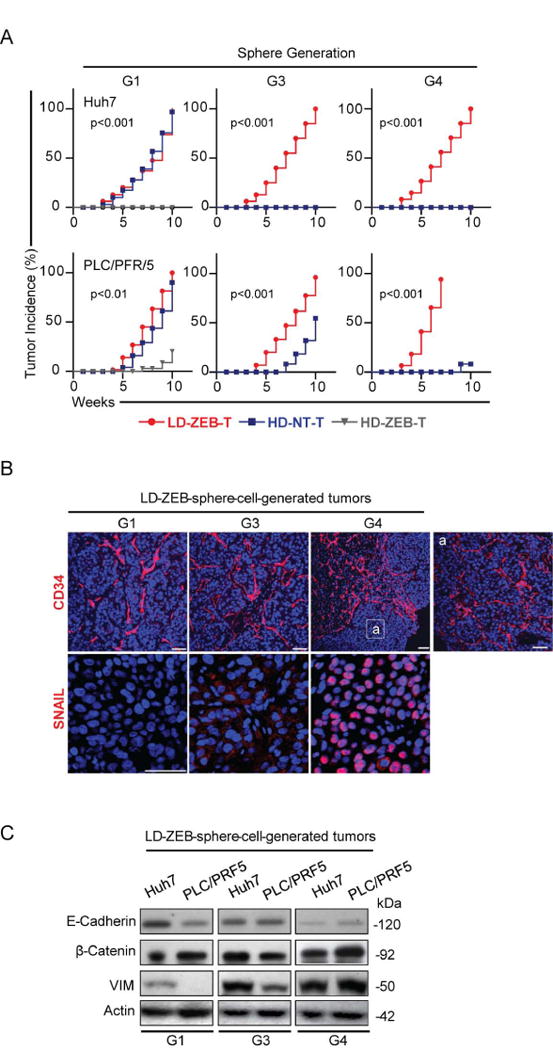
Reversal of DNA hypermethylation and associated gene silencing is an emerging cancer therapy approach. Here we addressed the impact of epigenetic alterations and cellular context on functional and transcriptional reprogramming of hepatocellular carcinoma (HCC) cells. Our strategy employed a 3-day treatment of established and primary human HCC-derived cell lines grown as a monolayer at various cell densities with the DNMT1 inhibitor zebularine (ZEB) followed by a 3D culture to identify cells endowed with self-renewal potential. Differences in self-renewal, gene expression, tumorigenicity, and metastatic potential of spheres at generations G1-G5 were examined. Transient ZEB exposure produced differential cell density-dependent responses. In cells grown at low density, ZEB caused a remarkable increase in self-renewal and tumorigenicity associated with long-lasting gene expression changes characterized by a stable overexpression of cancer stem cell-related and key epithelial-mesenchymal transition genes. These effects persisted after restoration of DNMT1 expression. In contrast, at high cell density, ZEB caused a gradual decrease in self-renewal and tumorigenicty, and up-regulation of apoptosis- and differentiation-related genes. A permanent reduction of DNMT1 protein using short hairpin RNA (shRNA)-mediated DNMT1 silencing rendered HCC cells insensitive both to cell density and ZEB effects. Similarly, WRL68 and HepG2 hepatoblastoma cells expressing low DNMT1 basal levels also possessed a high self-renewal, irrespective of cell density or ZEB exposure. Spheres formed by low-density cells treated with ZEB or shDNMT1 displayed a high molecular similarity which was sustained through consecutive generations, confirming the essential role of DNMT1 depletion in the enhancement of cancer stem cell properties.
Conclusion: These results identify DNA methylation as a key epigenetic regulatory mechanism determining the pool of cancer stem cells in liver cancer and possibly other solid tumors.
Published 2014. This article is a U.S. Government work and is in the public domain in the USA.
Figures
Similar articles
-
Osteopontin alters DNA methylation through up-regulating DNMT1 and sensitizes CD133+/CD44+ cancer stem cells to 5 azacytidine in hepatocellular carcinoma.J Exp Clin Cancer Res. 2018 Jul 31;37(1):179. doi: 10.1186/s13046-018-0832-1. J Exp Clin Cancer Res. 2018. PMID: 30064482 Free PMC article.
-
DNMT1-mediated methylation of BEX1 regulates stemness and tumorigenicity in liver cancer.J Hepatol. 2021 Nov;75(5):1142-1153. doi: 10.1016/j.jhep.2021.06.025. Epub 2021 Jul 1. J Hepatol. 2021. PMID: 34217777
-
PAF enhances cancer stem cell properties via β-catenin signaling in hepatocellular carcinoma.Cell Cycle. 2021 May;20(10):1010-1020. doi: 10.1080/15384101.2021.1919826. Epub 2021 May 10. Cell Cycle. 2021. PMID: 33970778 Free PMC article.
-
DNMT1 and DNMT3b silencing sensitizes human hepatoma cells to TRAIL-mediated apoptosis via up-regulation of TRAIL-R2/DR5 and caspase-8.Cancer Sci. 2010 Jun;101(6):1431-9. doi: 10.1111/j.1349-7006.2010.01565.x. Epub 2010 Mar 13. Cancer Sci. 2010. PMID: 20398055 Free PMC article.
-
Therapeutic approach of Cancer stem cells (CSCs) in gastric adenocarcinoma; DNA methyltransferases enzymes in cancer targeted therapy.Biomed Pharmacother. 2019 Jul;115:108958. doi: 10.1016/j.biopha.2019.108958. Epub 2019 May 7. Biomed Pharmacother. 2019. PMID: 31075731 Review.
Cited by
-
The Emerging Role of Tumor Microenvironmental Stimuli in Regulating Metabolic Rewiring of Liver Cancer Stem Cells.Cancers (Basel). 2022 Dec 20;15(1):5. doi: 10.3390/cancers15010005. Cancers (Basel). 2022. PMID: 36612000 Free PMC article. Review.
-
DNA Methyltransferases Modulate Hepatogenic Lineage Plasticity of Mesenchymal Stromal Cells.Stem Cell Reports. 2017 Jul 11;9(1):247-263. doi: 10.1016/j.stemcr.2017.05.008. Epub 2017 Jun 8. Stem Cell Reports. 2017. PMID: 28602611 Free PMC article.
-
Mechanism of cancer stemness maintenance in human liver cancer.Cell Death Dis. 2022 Apr 21;13(4):394. doi: 10.1038/s41419-022-04848-z. Cell Death Dis. 2022. PMID: 35449193 Free PMC article. Review.
-
Circulating MIR148A associates with sensitivity to adiponectin levels in human metabolic surgery for weight loss.Endocr Connect. 2018 Sep 1;7(9):975-982. doi: 10.1530/EC-18-0205. Endocr Connect. 2018. PMID: 30300537 Free PMC article.
-
Dysregulation of Iron Metabolism in Cholangiocarcinoma Stem-like Cells.Sci Rep. 2017 Dec 15;7(1):17667. doi: 10.1038/s41598-017-17804-1. Sci Rep. 2017. PMID: 29247214 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases