

Hoxb1b controls oriented cell division, cell shape and microtubule dynamics in neural tube morphogenesis
- PMID: 24449840
- PMCID: PMC3899817
- DOI: 10.1242/dev.098731
Hoxb1b controls oriented cell division, cell shape and microtubule dynamics in neural tube morphogenesis
Abstract
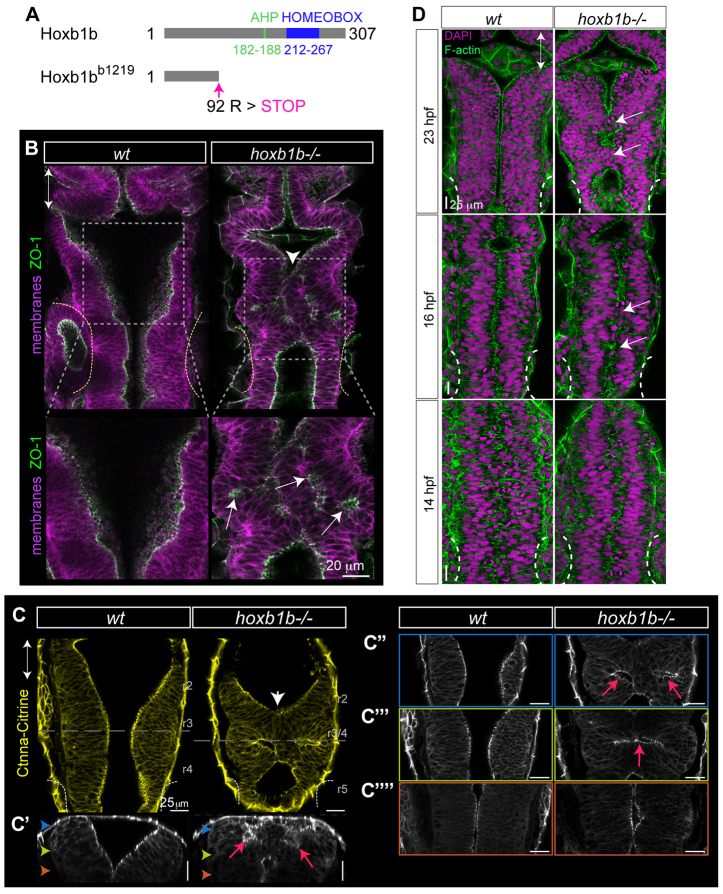
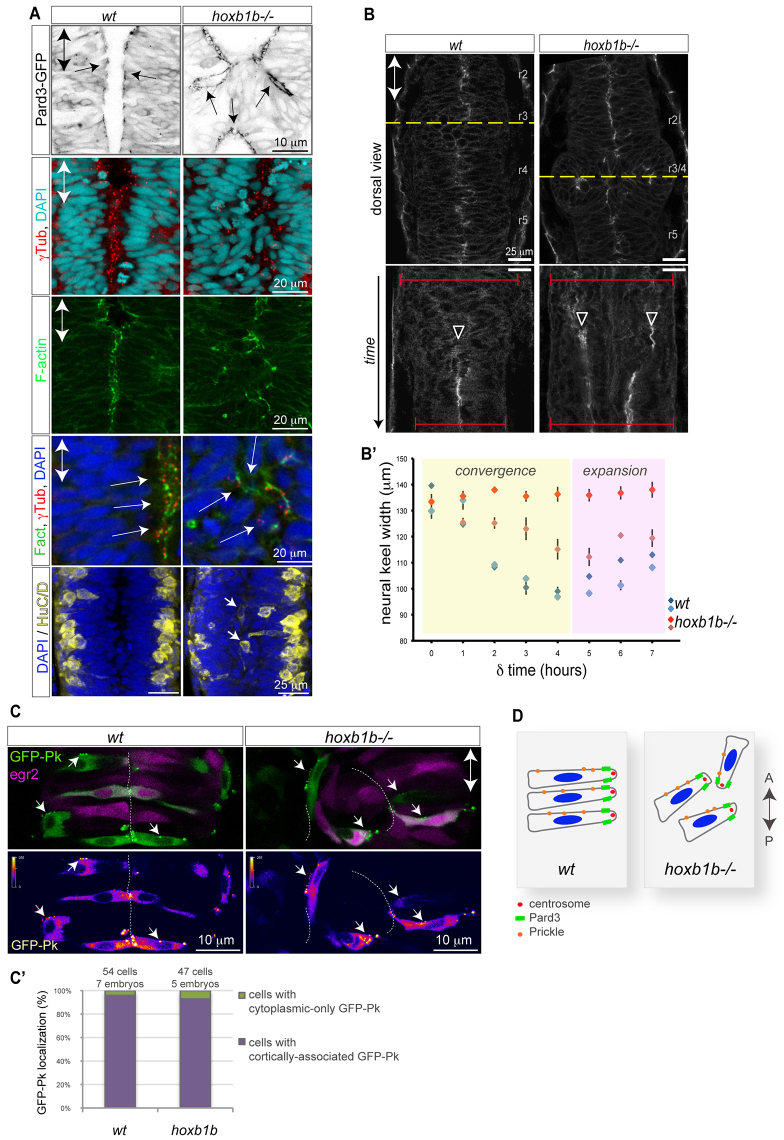
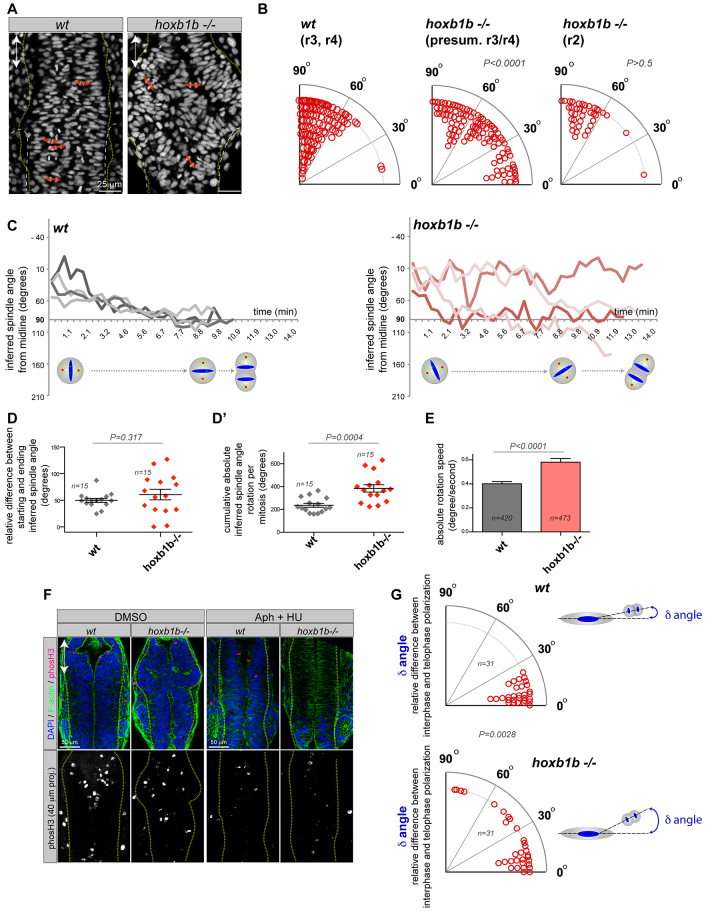
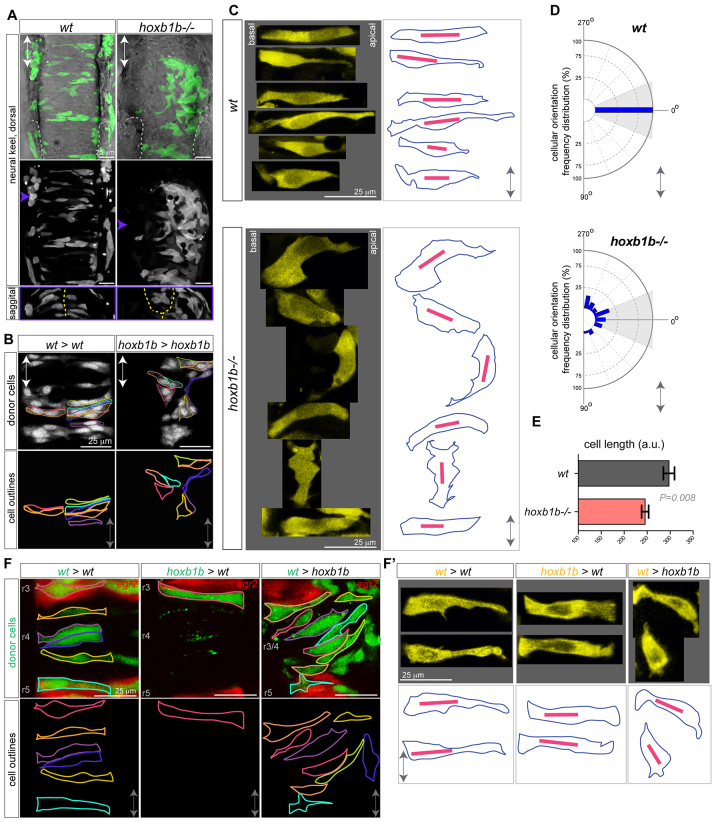
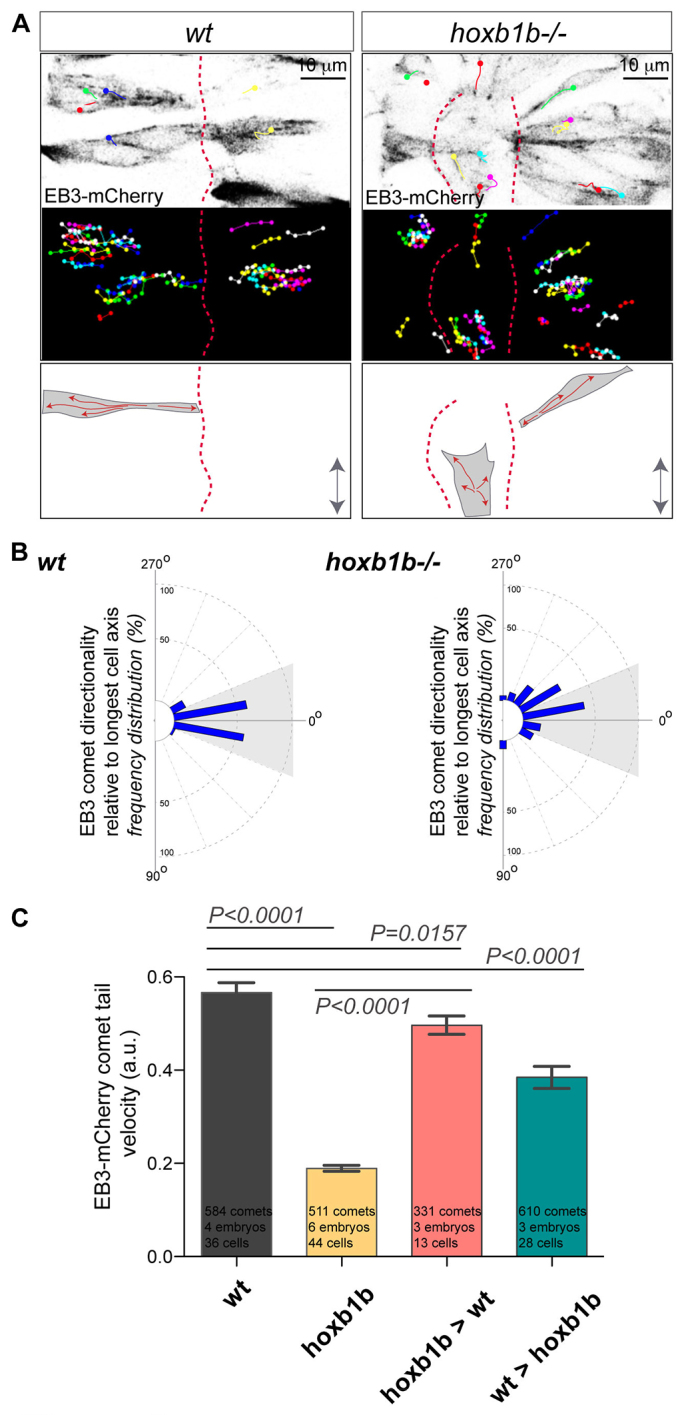
Hox genes are classically ascribed to function in patterning the anterior-posterior axis of bilaterian animals; however, their role in directing molecular mechanisms underlying morphogenesis at the cellular level remains largely unstudied. We unveil a non-classical role for the zebrafish hoxb1b gene, which shares ancestral functions with mammalian Hoxa1, in controlling progenitor cell shape and oriented cell division during zebrafish anterior hindbrain neural tube morphogenesis. This is likely distinct from its role in cell fate acquisition and segment boundary formation. We show that, without affecting major components of apico-basal or planar cell polarity, Hoxb1b regulates mitotic spindle rotation during the oriented neural keel symmetric mitoses that are required for normal neural tube lumen formation in the zebrafish. This function correlates with a non-cell-autonomous requirement for Hoxb1b in regulating microtubule plus-end dynamics in progenitor cells in interphase. We propose that Hox genes can influence global tissue morphogenesis by control of microtubule dynamics in individual cells in vivo.
Keywords: Cell polarity; Hoxa1; Hoxb1b; In vivo tissue morphogenesis; Neural tube formation; Oriented cell division; Zebrafish.
Figures
References
-
- Aubin J., Lemieux M., Tremblay M., Bérard J., Jeannotte L. (1997). Early postnatal lethality in Hoxa-5 mutant mice is attributable to respiratory tract defects. Dev. Biol. 192, 432–445 - PubMed
-
- Barrow J. R., Stadler H. S., Capecchi M. R. (2000). Roles of Hoxa1 and Hoxa2 in patterning the early hindbrain of the mouse. Development 127, 933–944 - PubMed
-
- Carpenter E. M., Goddard J. M., Chisaka O., Manley N. R., Capecchi M. R. (1993). Loss of Hox-A1 (Hox-1.6) function results in the reorganization of the murine hindbrain. Development 118, 1063–1075 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
