

Nonautonomous contact guidance signaling during collective cell migration
- PMID: 24449852
- PMCID: PMC3918762
- DOI: 10.1073/pnas.1321852111
Nonautonomous contact guidance signaling during collective cell migration
Abstract
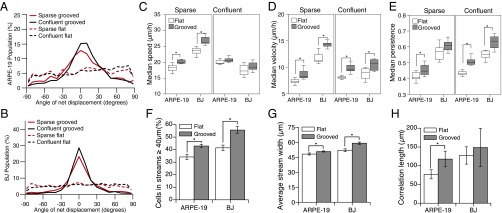
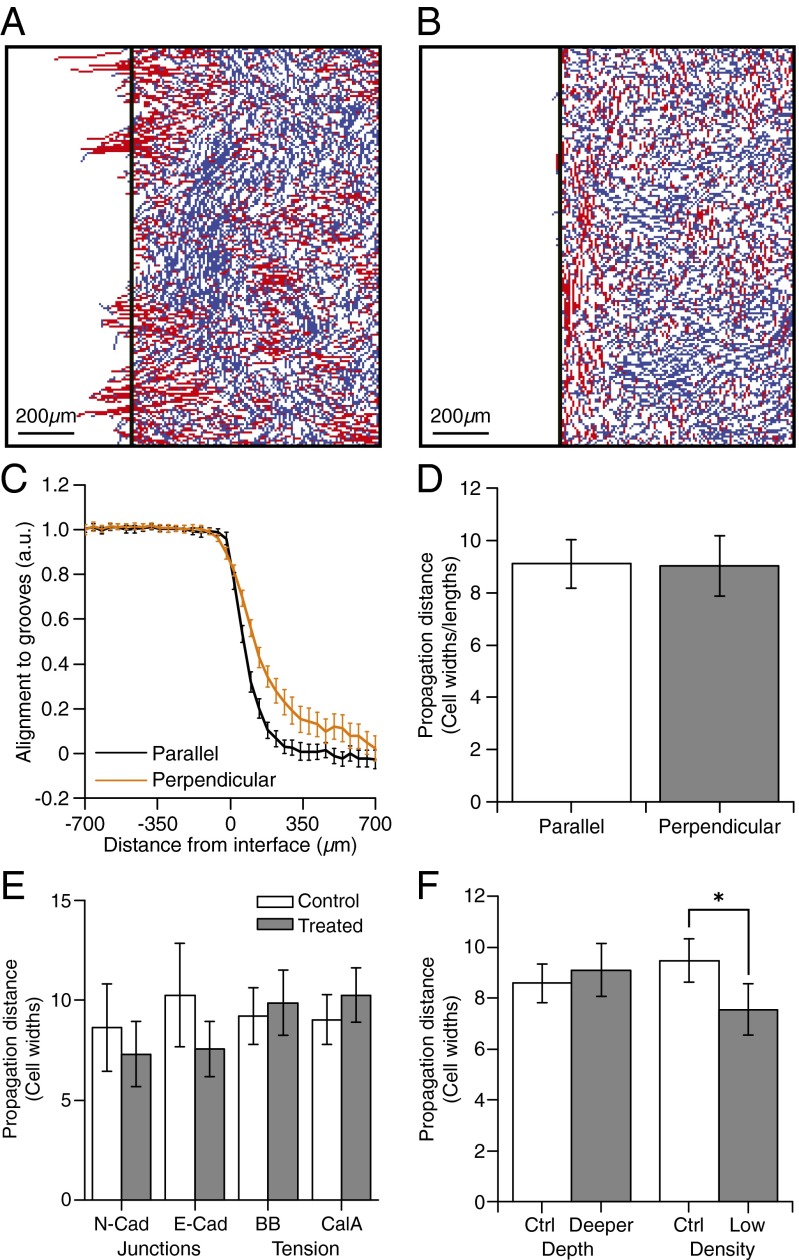
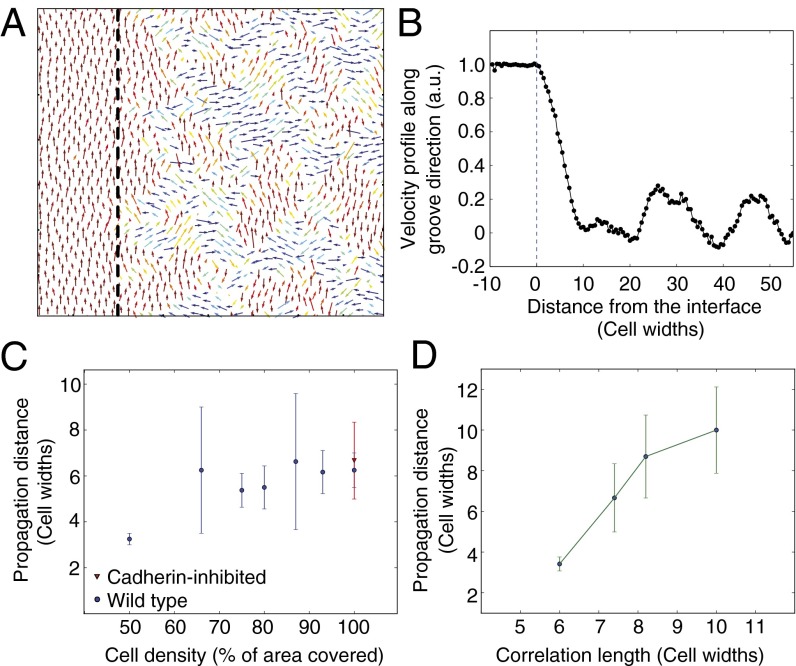
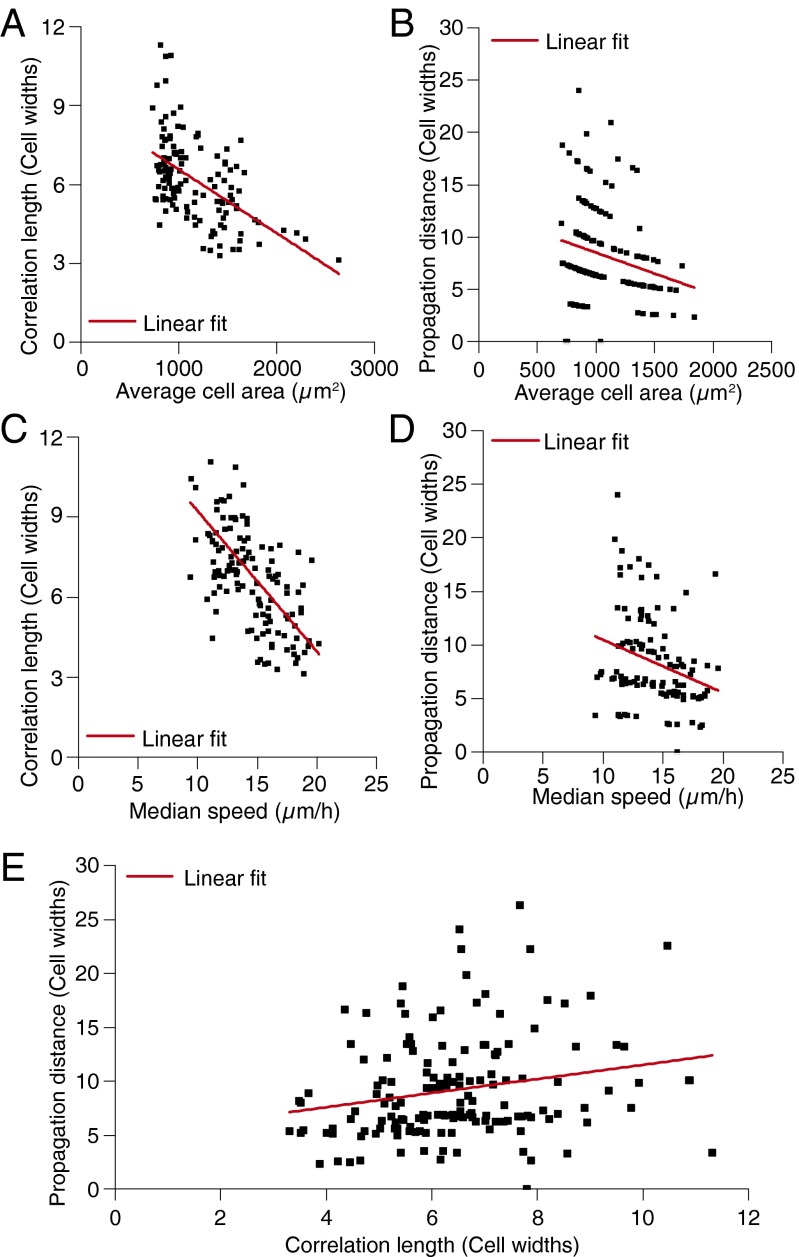
Directed migration of groups of cells is a critical aspect of tissue morphogenesis that ensures proper tissue organization and, consequently, function. Cells moving in groups, unlike single cells, must coordinate their migratory behavior to maintain tissue integrity. During directed migration, cells are guided by a combination of mechanical and chemical cues presented by neighboring cells and the surrounding extracellular matrix. One important class of signals that guide cell migration includes topographic cues. Although the contact guidance response of individual cells to topographic cues has been extensively characterized, little is known about the response of groups of cells to topographic cues, the impact of such cues on cell-cell coordination within groups, and the transmission of nonautonomous contact guidance information between neighboring cells. Here, we explore these phenomena by quantifying the migratory response of confluent monolayers of epithelial and fibroblast cells to contact guidance cues provided by grooved topography. We show that, in both sparse clusters and confluent sheets, individual cells are contact-guided by grooves and show more coordinated behavior on grooved versus flat substrates. Furthermore, we demonstrate both in vitro and in silico that the guidance signal provided by a groove can propagate between neighboring cells in a confluent monolayer, and that the distance over which signal propagation occurs is not significantly influenced by the strength of cell-cell junctions but is an emergent property, similar to cellular streaming, triggered by mechanical exclusion interactions within the collective system.
Keywords: correlation length; emergent behavior; group coordination; mechanical signal propagation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Friedl P, Gilmour D. Collective cell migration in morphogenesis, regeneration and cancer. Nat Rev Mol Cell Biol. 2009;10(7):445–457. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
