

Artificial riboswitches for gene expression and replication control of DNA and RNA viruses
- PMID: 24449891
- PMCID: PMC3918795
- DOI: 10.1073/pnas.1318563111
Artificial riboswitches for gene expression and replication control of DNA and RNA viruses
Abstract
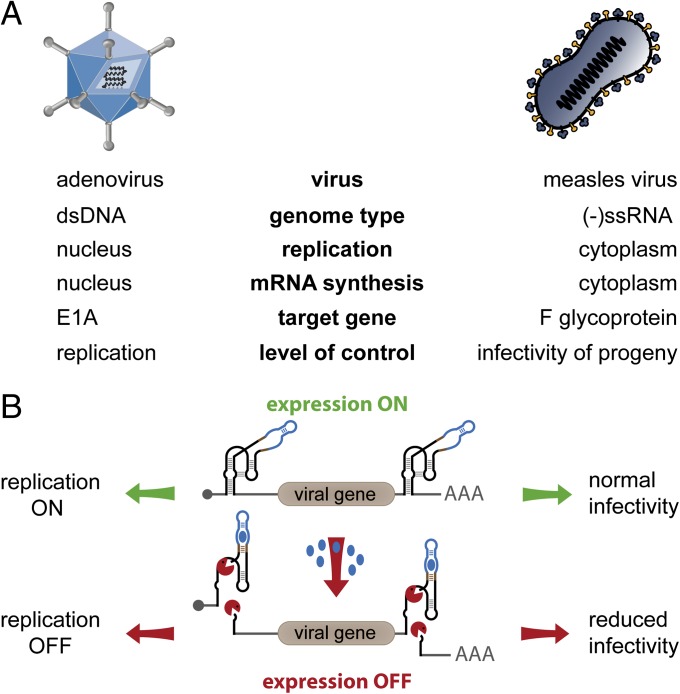
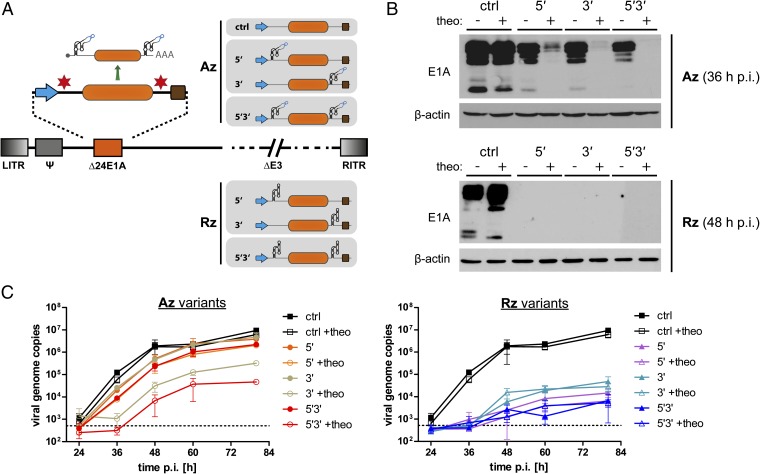
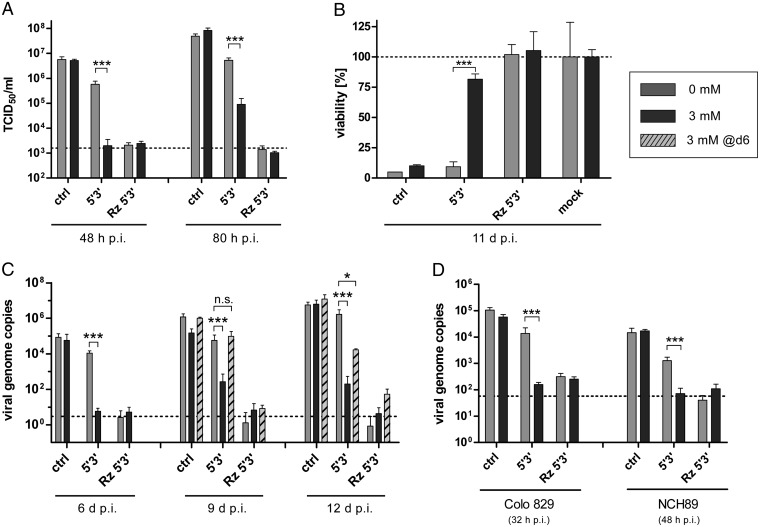
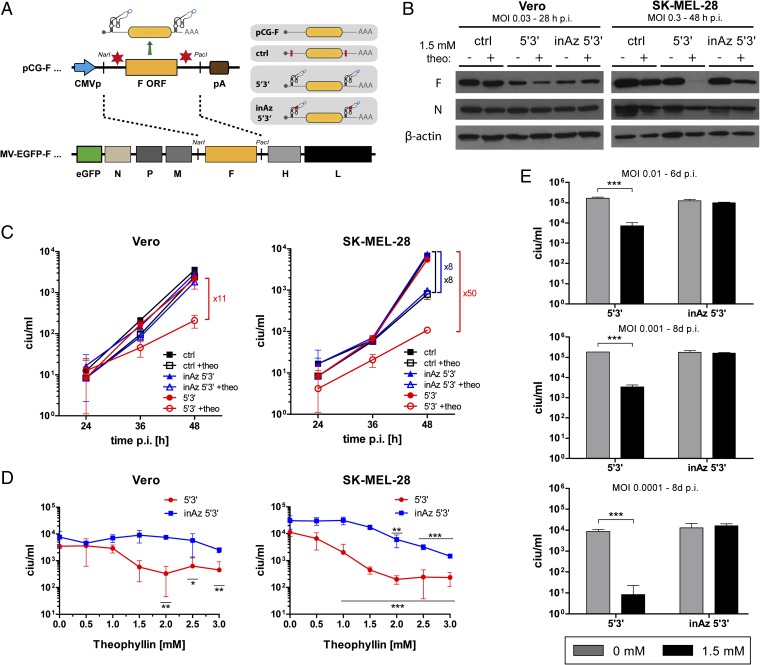
Aptazymes are small, ligand-dependent self-cleaving ribozymes that function independently of transcription factors and can be customized for induction by various small molecules. Here, we introduce these artificial riboswitches for regulation of DNA and RNA viruses. We hypothesize that they represent universally applicable tools for studying viral gene functions and for applications as a safety switch for oncolytic and live vaccine viruses. Our study shows that the insertion of artificial aptazymes into the adenoviral immediate early gene E1A enables small-molecule-triggered, dose-dependent inhibition of gene expression. Aptazyme-mediated shutdown of E1A expression translates into inhibition of adenoviral genome replication, infectious particle production, and cytotoxicity/oncolysis. These results provide proof of concept for the aptazyme approach for effective control of biological outcomes in eukaryotic systems, specifically in virus infections. Importantly, we also demonstrate aptazyme-dependent regulation of measles virus fusion protein expression, translating into potent reduction of progeny infectivity and virus spread. This not only establishes functionality of aptazymes in fully cytoplasmic genetic systems, but also implicates general feasibility of this strategy for application in viruses with either DNA or RNA genomes. Our study implies that gene regulation by artificial riboswitches may be an appealing alternative to Tet- and other protein-dependent gene regulation systems, based on their small size, RNA-intrinsic mode of action, and flexibility of the inducing molecule. Future applications range from gene analysis in basic research to medicine, for example as a safety switch for new generations of efficiency-enhanced oncolytic viruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
