

Spatiotemporal separation of PER and CRY posttranslational regulation in the mammalian circadian clock
- PMID: 24449901
- PMCID: PMC3918757
- DOI: 10.1073/pnas.1323618111
Spatiotemporal separation of PER and CRY posttranslational regulation in the mammalian circadian clock
Abstract
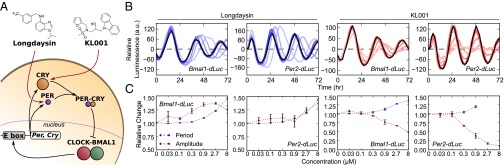
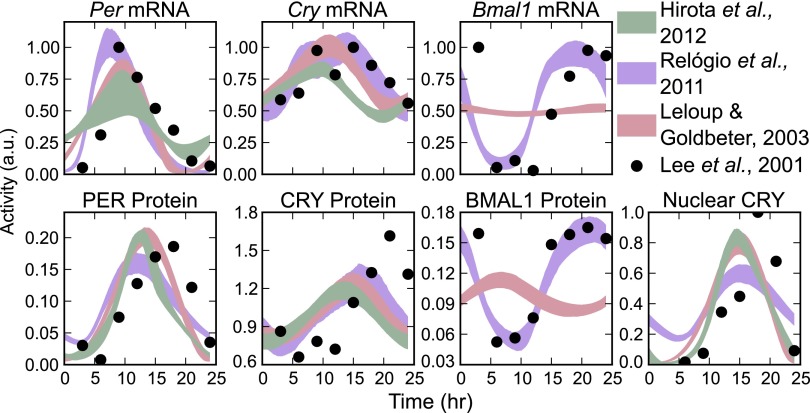
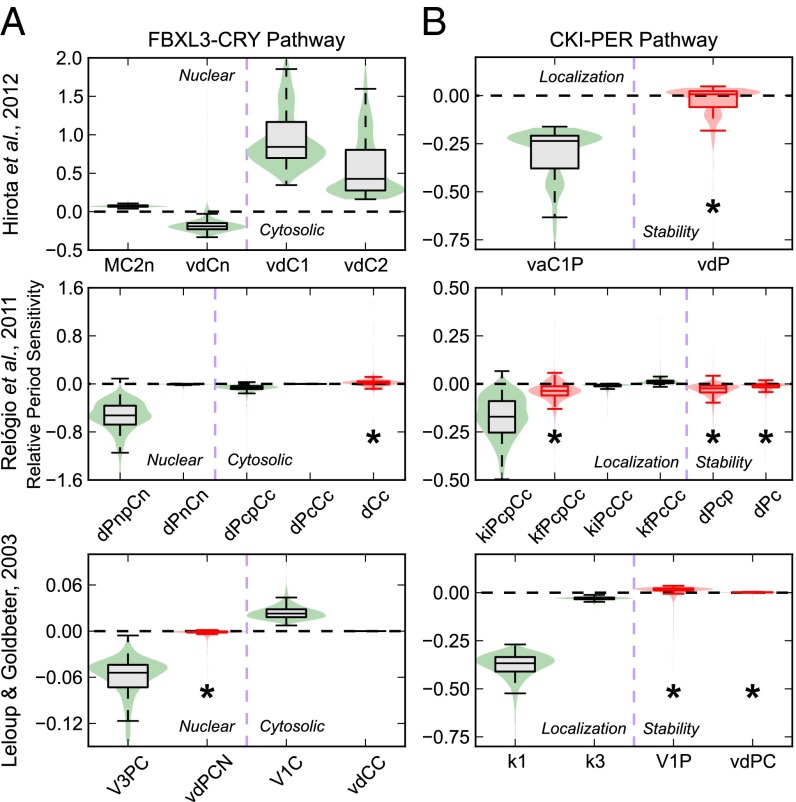
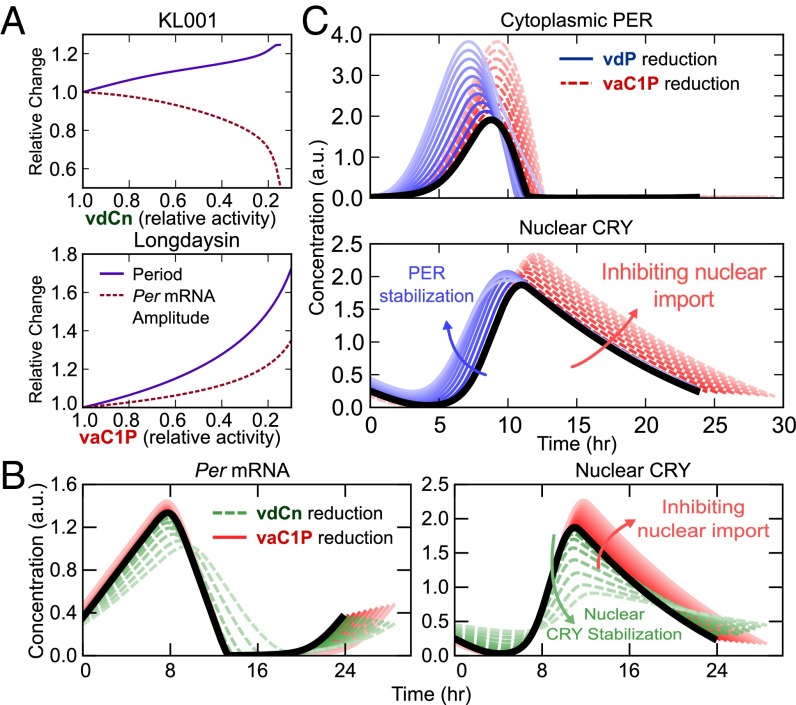
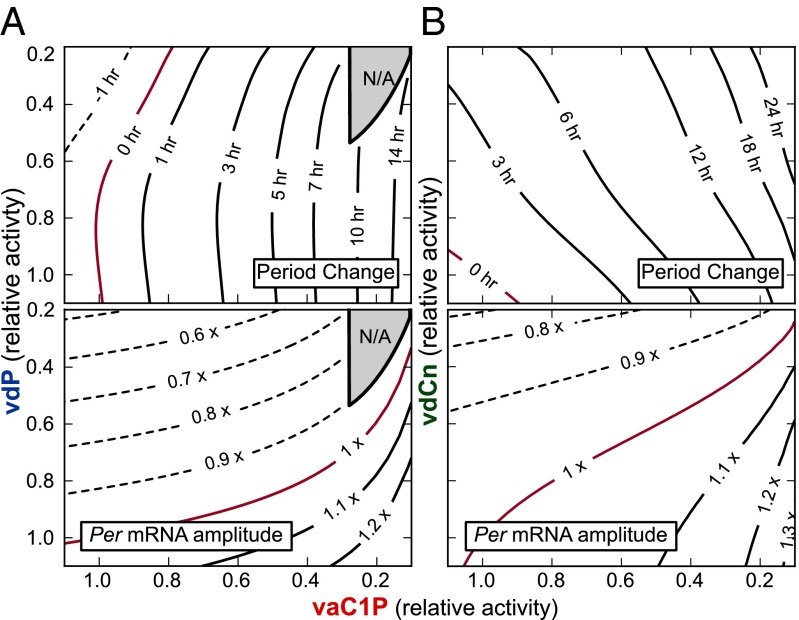
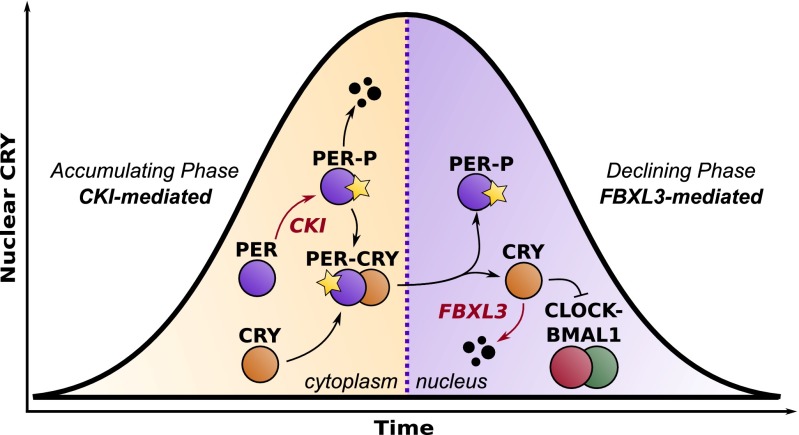
Posttranslational regulation of clock proteins is an essential part of mammalian circadian rhythms, conferring sensitivity to metabolic state and offering promising targets for pharmacological control. Two such regulators, casein kinase 1 (CKI) and F-box and leucine-rich repeat protein 3 (FBXL3), modulate the stability of closely linked core clock proteins period (PER) and cryptochrome (CRY), respectively. Inhibition of either CKI or FBXL3 leads to longer periods, and their effects are independent despite targeting proteins with similar roles in clock function. A mechanistic understanding of this independence, however, has remained elusive. Our analysis of cellular circadian clock gene reporters further differentiated between the actions of CKI and FBXL3 by revealing opposite amplitude responses from each manipulation. To understand the functional relationship between the CKI-PER and FBXL3-CRY pathways, we generated robust mechanistic predictions by applying a bootstrap uncertainty analysis to multiple mathematical circadian models. Our results indicate that CKI primarily regulates the accumulating phase of the PER-CRY repressive complex by controlling the nuclear import rate, whereas FBXL3 separately regulates the duration of transcriptional repression in the nucleus. Dynamic simulations confirmed that this spatiotemporal separation is able to reproduce the independence of the two regulators in period regulation, as well as their opposite amplitude effect. As a result, this study provides further insight into the molecular clock machinery responsible for maintaining robust circadian rhythms.
Keywords: gene regulation; identifiability analysis; sensitivity analysis.
Conflict of interest statement
Conflict of interest statement: S.A.K. is a scientific advisory board member of Reset Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
