

Structure and mechanism of a eukaryotic transmembrane ascorbate-dependent oxidoreductase
- PMID: 24449903
- PMCID: PMC3918761
- DOI: 10.1073/pnas.1323931111
Structure and mechanism of a eukaryotic transmembrane ascorbate-dependent oxidoreductase
Abstract
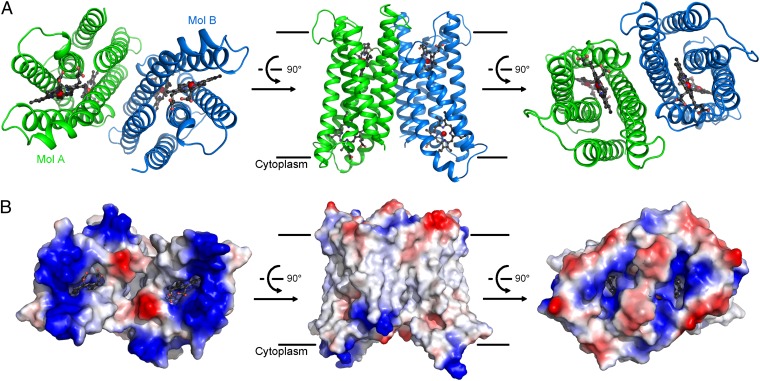
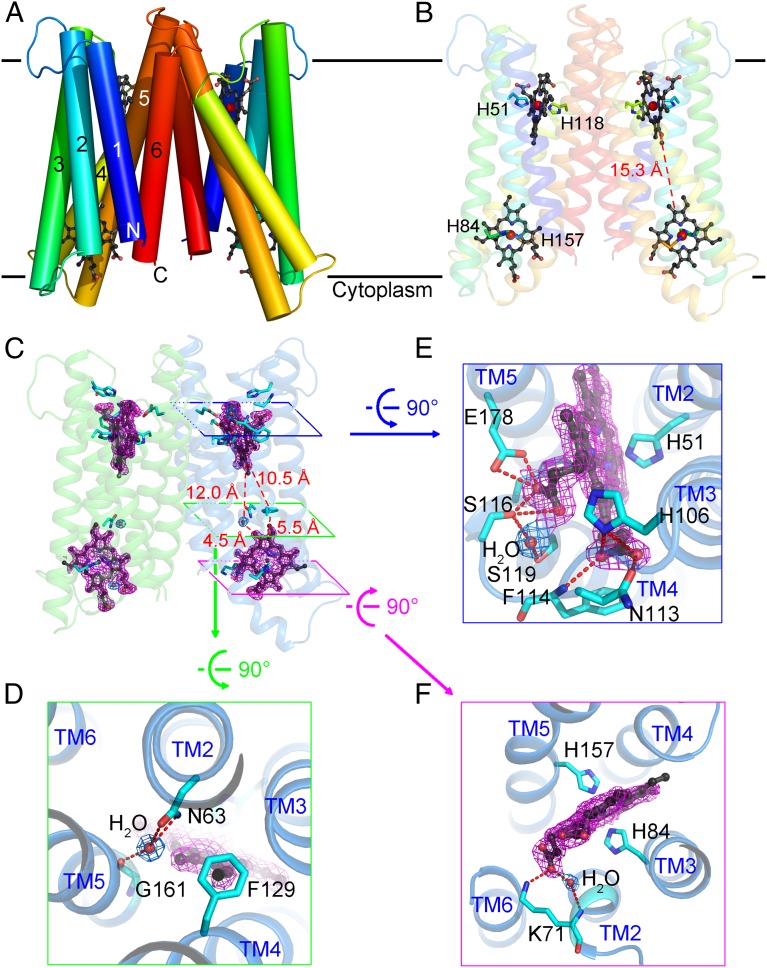
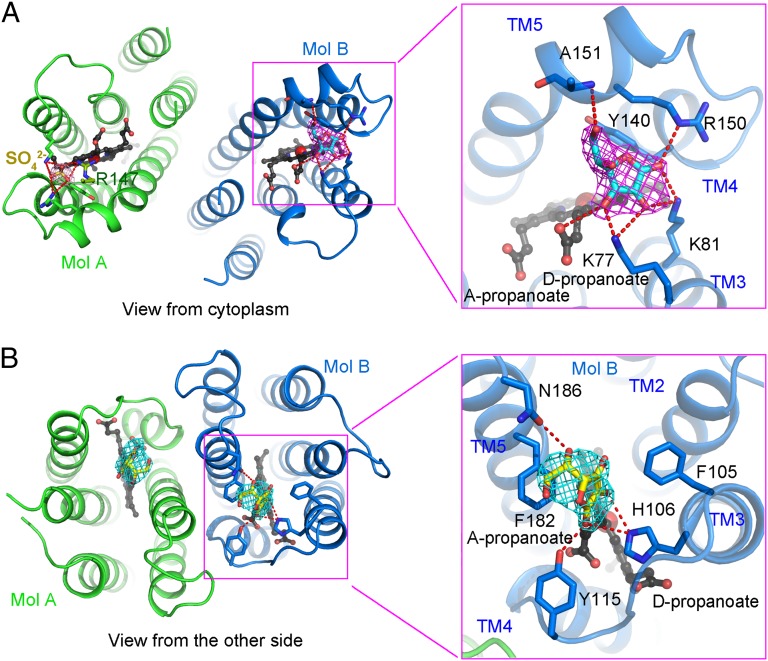
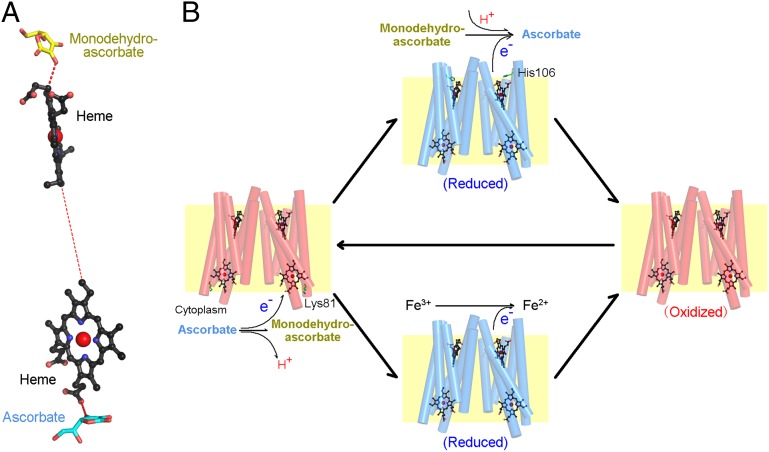
Vitamin C, also known as ascorbate, is required in numerous essential metabolic reactions in eukaryotes. The eukaryotic ascorbate-dependent oxidoreductase cytochrome b561 (Cyt b561), a family of highly conserved transmembrane enzymes, plays an important role in ascorbate recycling and iron absorption. Although Cyt b561 was identified four decades ago, its atomic structure and functional mechanism remain largely unknown. Here, we report the high-resolution crystal structures of cytochrome b561 from Arabidopsis thaliana in both substrate-free and substrate-bound states. Cyt b561 forms a homodimer, with each protomer consisting of six transmembrane helices and two heme groups. The negatively charged substrate ascorbate, or monodehydroascorbate, is enclosed in a positively charged pocket on either side of the membrane. Two highly conserved amino acids, Lys(81) and His(106), play an essential role in substrate recognition and catalysis. Our structural and biochemical analyses allow the proposition of a general electron transfer mechanism for members of the Cyt b561 family.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Roles of conserved Arg(72) and Tyr(71) in the ascorbate-specific transmembrane electron transfer catalyzed by Zea mays cytochrome b561.J Biosci Bioeng. 2013 May;115(5):497-506. doi: 10.1016/j.jbiosc.2012.11.013. Epub 2013 Jan 3. J Biosci Bioeng. 2013. PMID: 23290447
-
Higher-plant plasma membrane cytochrome b561: a protein in search of a function.Protoplasma. 2001;217(1-3):77-93. doi: 10.1007/BF01289417. Protoplasma. 2001. PMID: 11732342 Review.
-
Identification of an ascorbate-dependent cytochrome b of the tonoplast membrane sharing biochemical features with members of the cytochrome b561 family.Planta. 2005 Jan;220(3):365-75. doi: 10.1007/s00425-004-1360-0. Epub 2004 Sep 9. Planta. 2005. PMID: 15365836
-
Histidine cycle mechanism for the concerted proton/electron transfer from ascorbate to the cytosolic haem b centre of cytochrome b561: a unique machinery for the biological transmembrane electron transfer.J Biochem. 2007 Nov;142(5):553-60. doi: 10.1093/jb/mvm181. Epub 2007 Sep 28. J Biochem. 2007. PMID: 17905810
-
Cytochrome b561, ascorbic acid, and transmembrane electron transfer.Am J Clin Nutr. 1991 Dec;54(6 Suppl):1173S-1178S. doi: 10.1093/ajcn/54.6.1173s. Am J Clin Nutr. 1991. PMID: 1962566 Review.
Cited by
-
Duodenal cytochrome b (DCYTB) in iron metabolism: an update on function and regulation.Nutrients. 2015 Mar 31;7(4):2274-96. doi: 10.3390/nu7042274. Nutrients. 2015. PMID: 25835049 Free PMC article. Review.
-
Comparison of insect and human cytochrome b561 proteins: Insights into candidate ferric reductases in insects.PLoS One. 2023 Dec 1;18(12):e0291564. doi: 10.1371/journal.pone.0291564. eCollection 2023. PLoS One. 2023. PMID: 38039324 Free PMC article.
-
Induced Resistance Mechanism of Novel Curcumin Analogs Bearing a Quinazoline Moiety to Plant Virus.Int J Mol Sci. 2018 Dec 15;19(12):4065. doi: 10.3390/ijms19124065. Int J Mol Sci. 2018. PMID: 30558295 Free PMC article.
-
The recombinant expression systems for structure determination of eukaryotic membrane proteins.Protein Cell. 2014 Sep;5(9):658-72. doi: 10.1007/s13238-014-0086-4. Epub 2014 Aug 15. Protein Cell. 2014. PMID: 25119489 Free PMC article. Review.
-
Manipulating the position of DNA expression cassettes using location tags fused to dCas9 (Cas9-Lag) to improve metabolic pathway efficiency.Microb Cell Fact. 2020 Dec 14;19(1):229. doi: 10.1186/s12934-020-01496-w. Microb Cell Fact. 2020. PMID: 33317552 Free PMC article.
References
-
- Zetterström R. Nobel Prize 1937 to Albert von Szent-Györgyi: Identification of vitamin C as the anti-scorbutic factor. Acta Paediatr. 2009;98(5):915–919. - PubMed
-
- Peterkofsky B. Ascorbate requirement for hydroxylation and secretion of procollagen: Relationship to inhibition of collagen synthesis in scurvy. Am J Clin Nutr. 1991;54(6) Suppl:1135S–1140S. - PubMed
-
- Rebouche CJ. Ascorbic acid and carnitine biosynthesis. Am J Clin Nutr. 1991;54(6) Suppl:1147S–1152S. - PubMed
-
- Kaufman S. Dopamine-beta-hydroxylase. J Psychiatr Res. 1974;11:303–316. - PubMed
-
- Padayatty SJ, et al. Vitamin C as an antioxidant: Evaluation of its role in disease prevention. J Am Coll Nutr. 2003;22(1):18–35. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
