

Essential role of poly(ADP-ribosyl)ation in cocaine action
- PMID: 24449909
- PMCID: PMC3918779
- DOI: 10.1073/pnas.1319703111
Essential role of poly(ADP-ribosyl)ation in cocaine action
Abstract
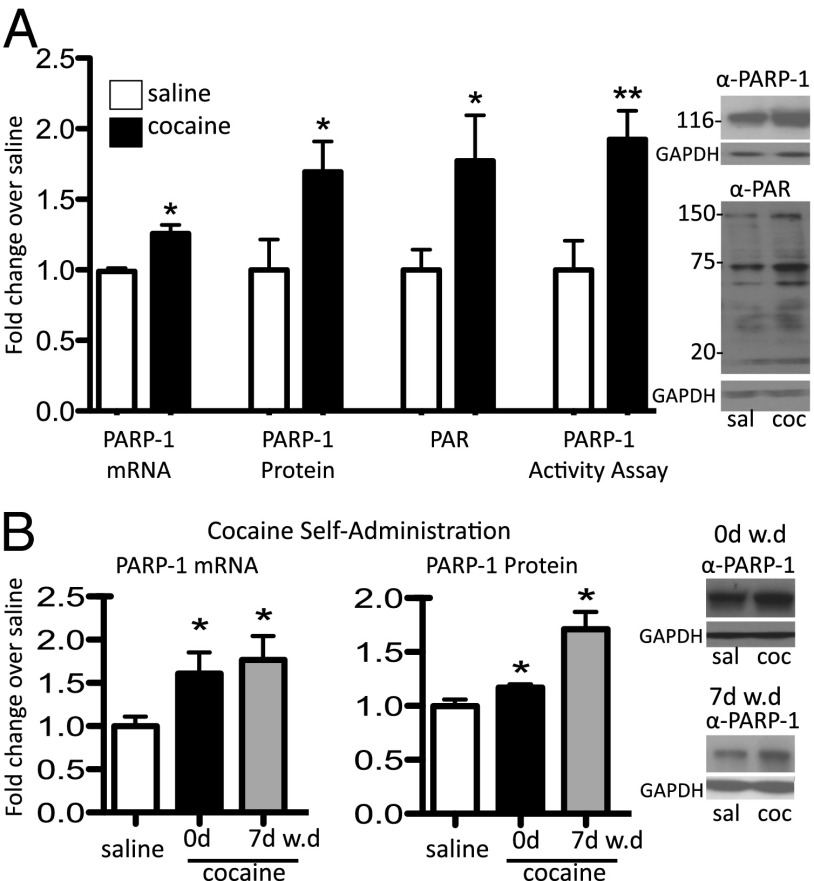
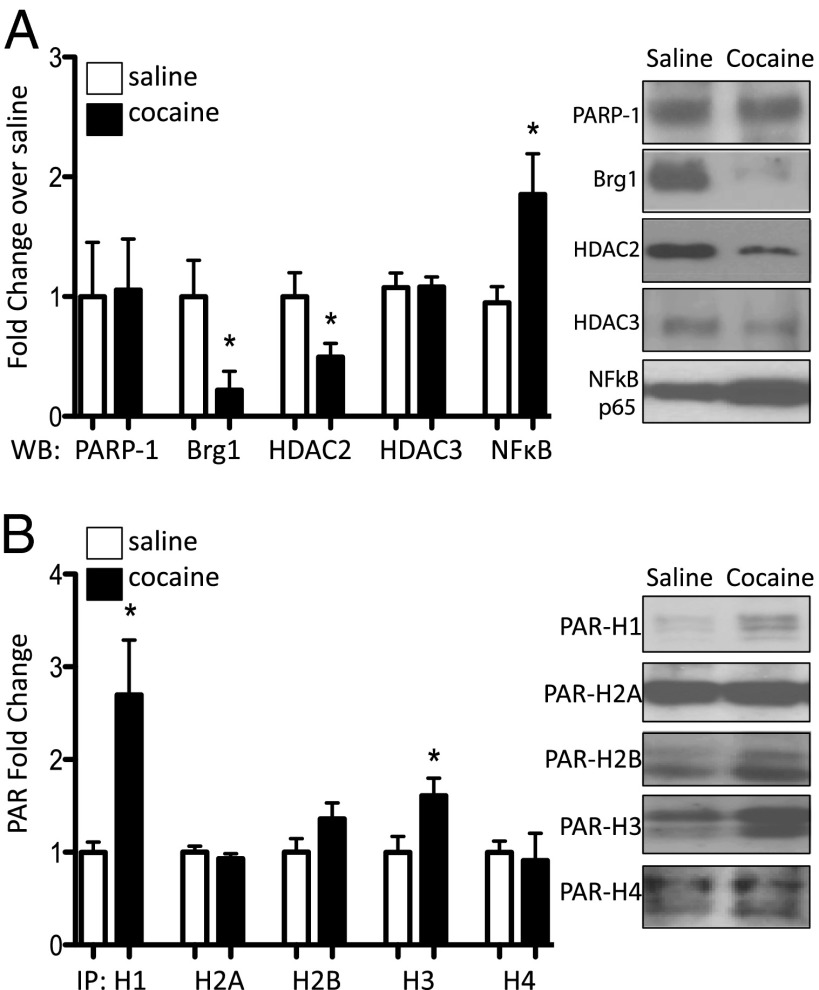
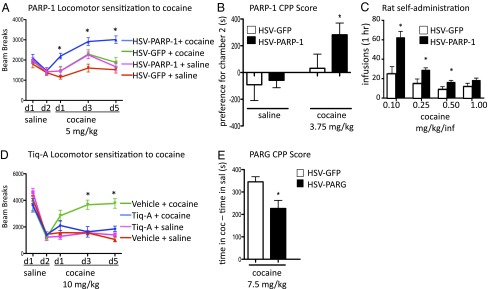
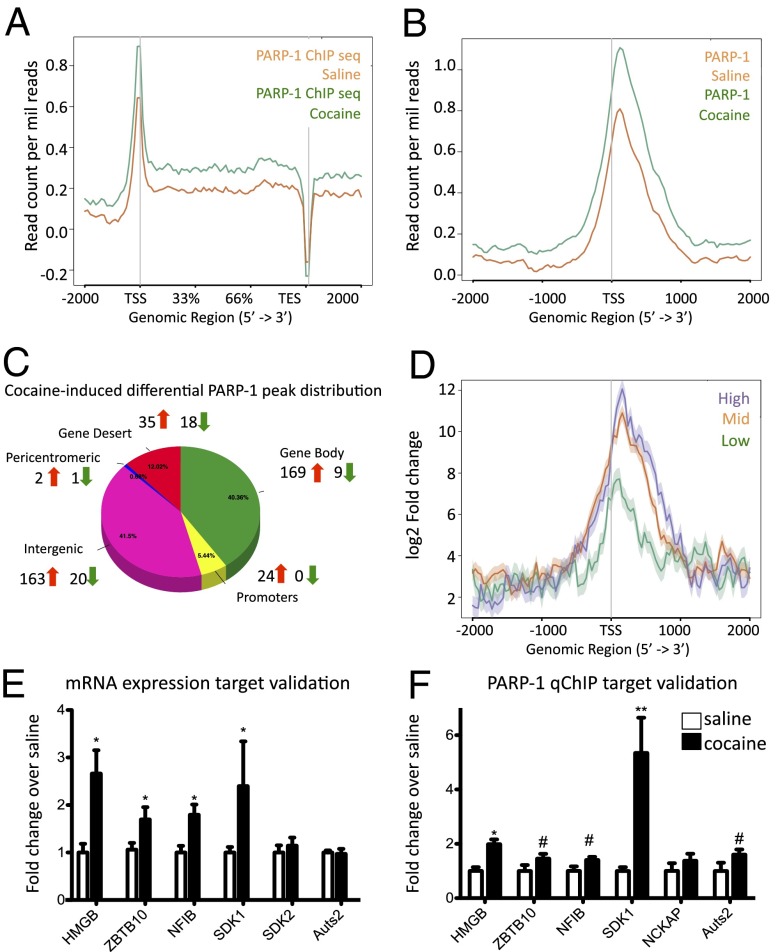
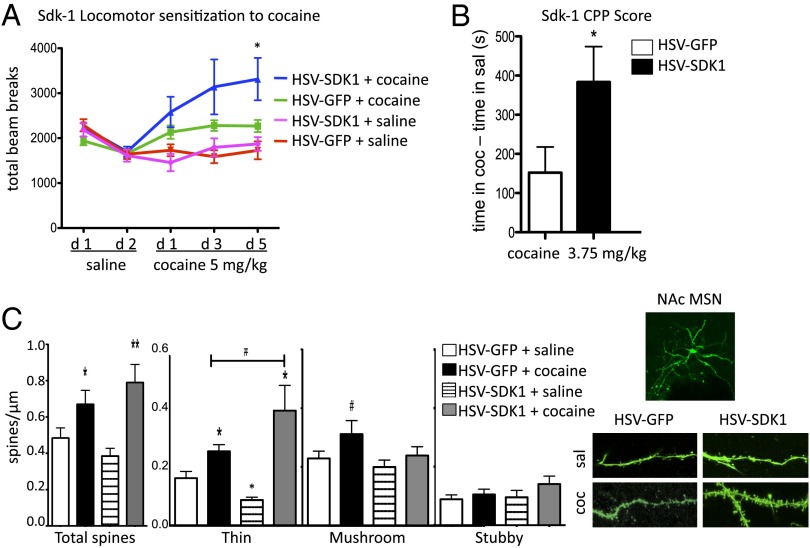
Many of the long-term effects of cocaine on the brain's reward circuitry have been shown to be mediated by alterations in gene expression. Several chromatin modifications, including histone acetylation and methylation, have been implicated in this regulation, but the effect of other histone modifications remains poorly understood. Poly(ADP-ribose) polymerase-1 (PARP-1), a ubiquitous and abundant nuclear protein, catalyzes the synthesis of a negatively charged polymer called poly(ADP-ribose) or PAR on histones and other substrate proteins and forms transcriptional regulatory complexes with several other chromatin proteins. Here, we identify an essential role for PARP-1 in cocaine-induced molecular, neural, and behavioral plasticity. Repeated cocaine administration, including self-administration, increased global levels of PARP-1 and its mark PAR in mouse nucleus accumbens (NAc), a key brain reward region. Using PARP-1 inhibitors and viral-mediated gene transfer, we established that PARP-1 induction in NAc mediates enhanced behavioral responses to cocaine, including increased self-administration of the drug. Using chromatin immunoprecipitation sequencing, we demonstrated a global, genome-wide enrichment of PARP-1 in NAc of cocaine-exposed mice and identified several PARP-1 target genes that could contribute to the lasting effects of cocaine. Specifically, we identified sidekick-1--important for synaptic connections during development--as a critical PARP-1 target gene involved in cocaine's behavioral effects as well as in its ability to induce dendritic spines on NAc neurons. These findings establish the involvement of PARP-1 and PARylation in the long-term actions of cocaine.
Keywords: drug addiction; histone PARylation; medium spiny neurons.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
