

Structural basis for nuclear import of splicing factors by human Transportin 3
- PMID: 24449914
- PMCID: PMC3932936
- DOI: 10.1073/pnas.1320755111
Structural basis for nuclear import of splicing factors by human Transportin 3
Abstract
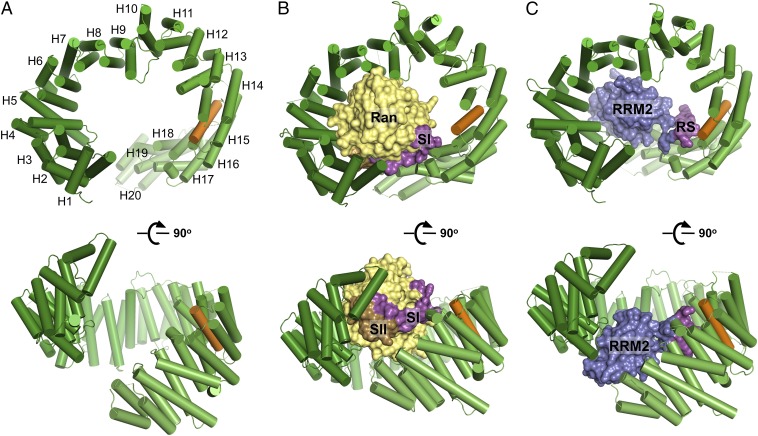
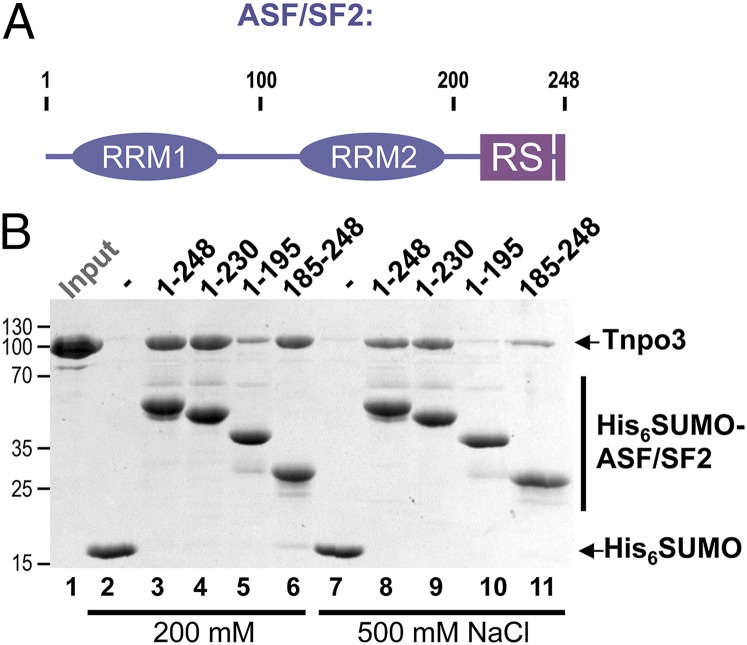
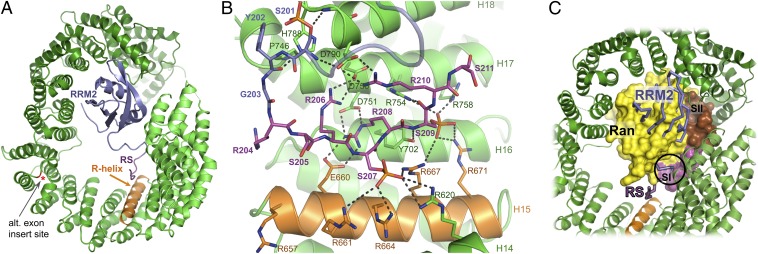
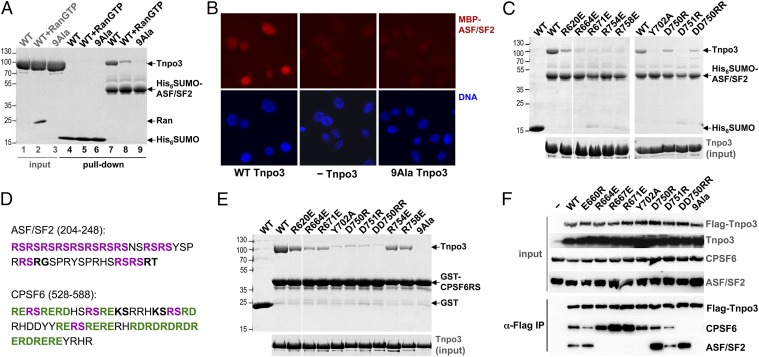
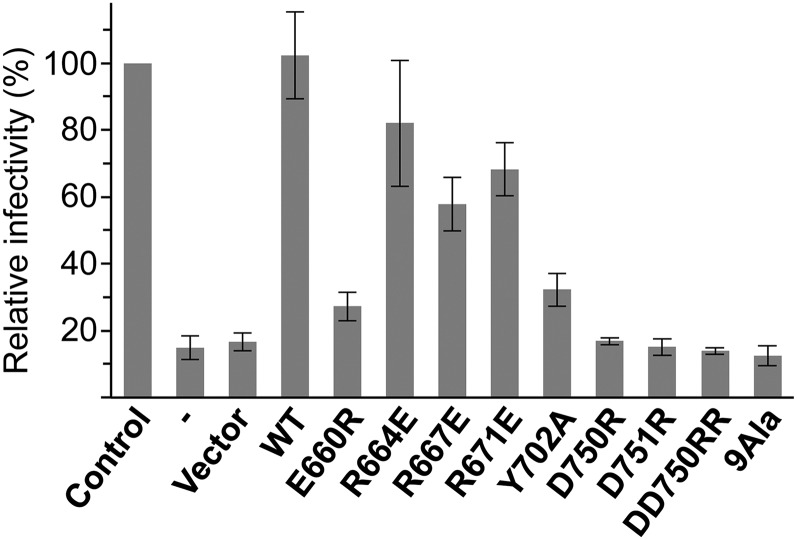
Transportin 3 (Tnpo3, Transportin-SR2) is implicated in nuclear import of splicing factors and HIV-1 replication. Herein, we show that the majority of cellular Tnpo3 binding partners contain arginine-serine (RS) repeat domains and present crystal structures of human Tnpo3 in its free as well as GTPase Ran- and alternative splicing factor/splicing factor 2 (ASF/SF2)-bound forms. The flexible β-karyopherin fold of Tnpo3 embraces the RNA recognition motif and RS domains of the cargo. A constellation of charged residues on and around the arginine-rich helix of Tnpo3 HEAT repeat 15 engage the phosphorylated RS domain and are critical for the recognition and nuclear import of ASF/SF2. Mutations in the same region of Tnpo3 impair its interaction with the cleavage and polyadenylation specificity factor 6 (CPSF6) and its ability to support HIV-1 replication. Steric incompatibility of the RS domain and RanGTP engagement by Tnpo3 provides the mechanism for cargo release in the nucleus. Our results elucidate the structural bases for nuclear import of splicing factors and the Tnpo3-CPSF6 nexus in HIV-1 biology.
Keywords: SR protein; Transportin-SR; host factor; importin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Stewart M. Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol. 2007;8(3):195–208. - PubMed
-
- Cook A, Bono F, Jinek M, Conti E. Structural biology of nucleocytoplasmic transport. Annu Rev Biochem. 2007;76:647–671. - PubMed
-
- Cook AG, Conti E. Nuclear export complexes in the frame. Curr Opin Struct Biol. 2010;20(2):247–252. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
