

PIKE is essential for oligodendroglia development and CNS myelination
- PMID: 24449917
- PMCID: PMC3918755
- DOI: 10.1073/pnas.1318185111
PIKE is essential for oligodendroglia development and CNS myelination
Abstract
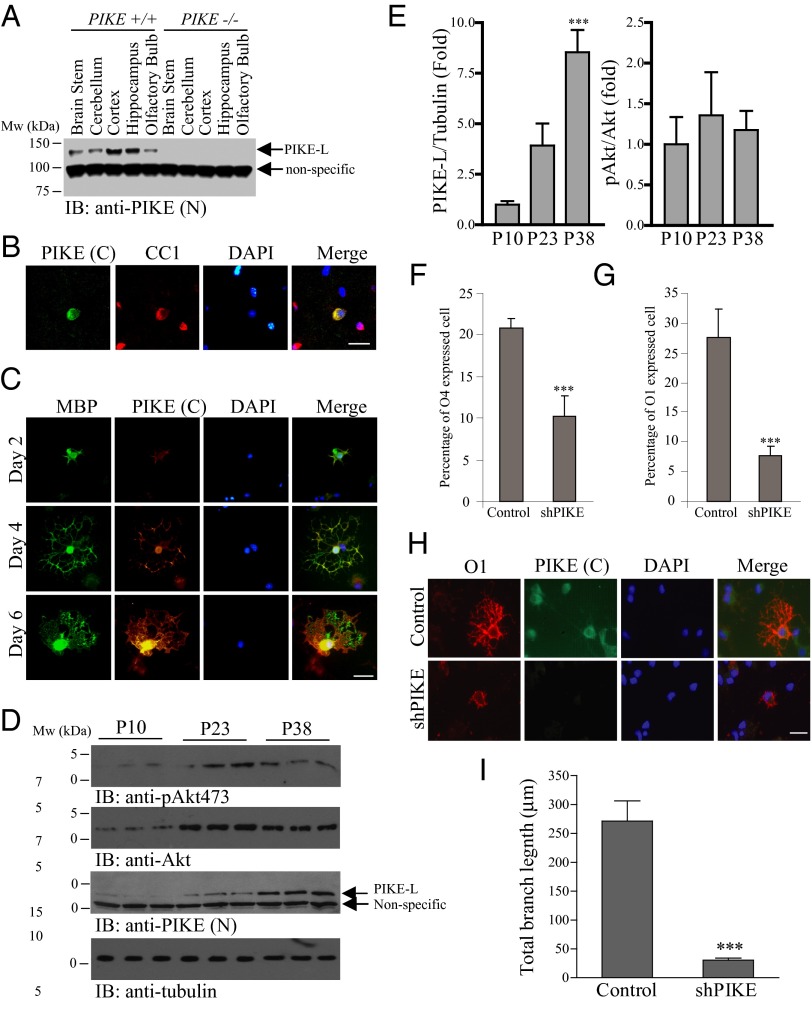
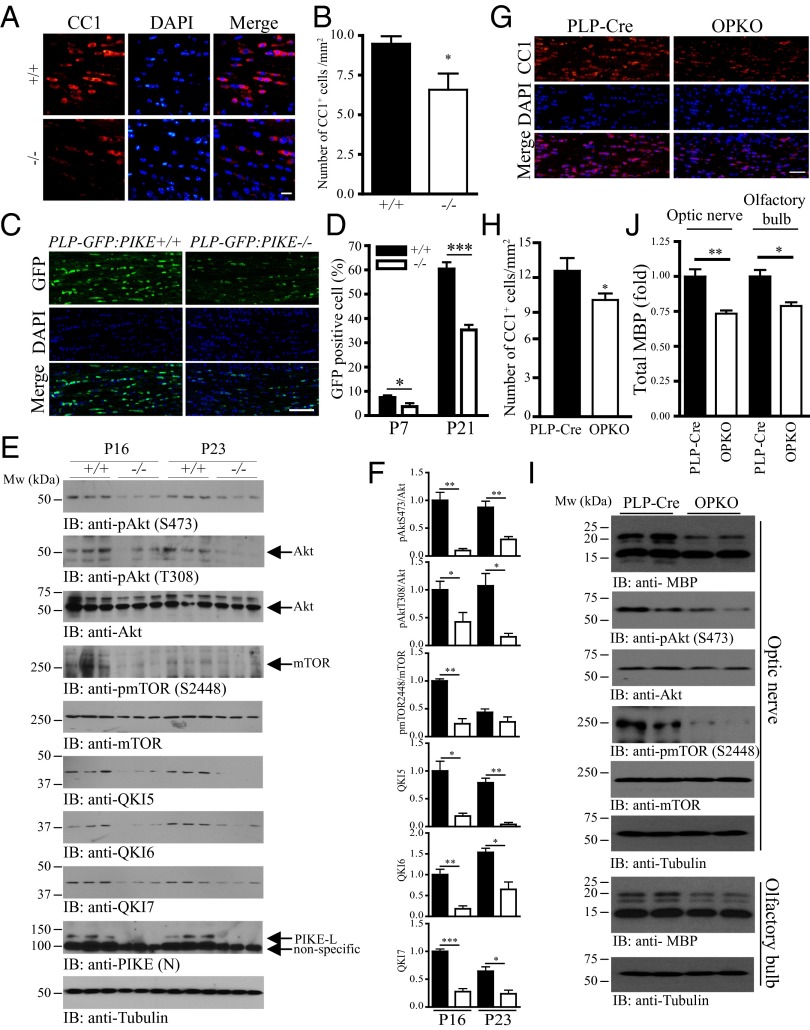
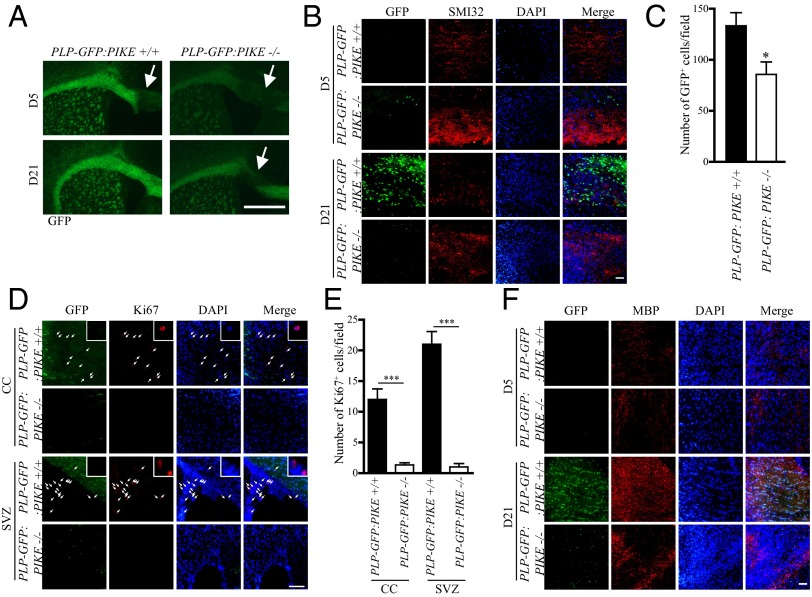
Oligodendrocyte (OL) differentiation and myelin development are complex events regulated by numerous signal transduction factors. Here, we report that phosphoinositide-3 kinase enhancer L (PIKE-L) is required for OL development and myelination. PIKE-L expression is up-regulated when oligodendrocyte progenitor cells commit to differentiation. Conversely, depleting phosphoinositide-3 kinase enhancer (PIKE) expression by shRNA prevents oligodendrocyte progenitor cell differentiation. In both conventional PIKE knockout (PIKE(-/-)) and OL-specific PIKE knockout mice, the number of OLs is reduced in the corpus callosum. PIKE(-/-) OLs also display defects when forming myelin sheath on neuronal axons during neonatal development, which is partially rescued when PTEN is ablated. In addition, Akt/mTOR signaling is impaired in OL-enriched tissues of the PIKE(-/-) mutant, leading to reduced expression of critical proteins for myelin development and hypomyelination. Moreover, myelin repair of lysolecithin-induced lesions is delayed in PIKE(-/-) brain. Thus, PIKE plays pivotal roles to advance OL development and myelinogenesis through Akt/mTOR activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
R-Ras1 and R-Ras2 Are Essential for Oligodendrocyte Differentiation and Survival for Correct Myelination in the Central Nervous System.J Neurosci. 2018 May 30;38(22):5096-5110. doi: 10.1523/JNEUROSCI.3364-17.2018. Epub 2018 May 2. J Neurosci. 2018. PMID: 29720552 Free PMC article.
-
Mammalian target of rapamycin promotes oligodendrocyte differentiation, initiation and extent of CNS myelination.J Neurosci. 2014 Mar 26;34(13):4453-65. doi: 10.1523/JNEUROSCI.4311-13.2014. J Neurosci. 2014. PMID: 24671992 Free PMC article.
-
mTORC2 Loss in Oligodendrocyte Progenitor Cells Results in Regional Hypomyelination in the Central Nervous System.J Neurosci. 2023 Jan 25;43(4):540-558. doi: 10.1523/JNEUROSCI.0010-22.2022. Epub 2022 Dec 2. J Neurosci. 2023. PMID: 36460463 Free PMC article.
-
Nudging oligodendrocyte intrinsic signaling to remyelinate and repair: Estrogen receptor ligand effects.J Steroid Biochem Mol Biol. 2016 Jun;160:43-52. doi: 10.1016/j.jsbmb.2016.01.006. Epub 2016 Jan 14. J Steroid Biochem Mol Biol. 2016. PMID: 26776441 Free PMC article. Review.
-
Akt/mTOR signalling in myelination.Biochem Soc Trans. 2013 Aug;41(4):944-50. doi: 10.1042/BST20130046. Biochem Soc Trans. 2013. PMID: 23863161 Review.
Cited by
-
Septin/anillin filaments scaffold central nervous system myelin to accelerate nerve conduction.Elife. 2016 Aug 9;5:e17119. doi: 10.7554/eLife.17119. Elife. 2016. PMID: 27504968 Free PMC article.
-
Activation of the Akt Attenuates Ropivacaine-Induced Myelination Impairment in Spinal Cord and Sensory Dysfunction in Neonatal Rats.Mol Neurobiol. 2023 Dec;60(12):7009-7020. doi: 10.1007/s12035-023-03498-2. Epub 2023 Jul 31. Mol Neurobiol. 2023. PMID: 37523045
-
AGAP duplicons associate with structural diversity at Chromosome 10q11.22.Genome Res. 2024 Oct 29;34(10):1487-1499. doi: 10.1101/gr.279454.124. Genome Res. 2024. PMID: 39322278 Free PMC article.
-
α-Synuclein binds and sequesters PIKE-L into Lewy bodies, triggering dopaminergic cell death via AMPK hyperactivation.Proc Natl Acad Sci U S A. 2017 Jan 31;114(5):1183-1188. doi: 10.1073/pnas.1618627114. Epub 2017 Jan 17. Proc Natl Acad Sci U S A. 2017. PMID: 28096359 Free PMC article.
-
RIT1 regulation of CNS lipids RIT1 deficiency Alters cerebral lipid metabolism and reduces white matter tract oligodendrocytes and conduction velocities.Heliyon. 2023 Sep 23;9(10):e20384. doi: 10.1016/j.heliyon.2023.e20384. eCollection 2023 Oct. Heliyon. 2023. PMID: 37780758 Free PMC article.
References
-
- Baumann N, Pham-Dinh D. Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiol Rev. 2001;81(2):871–927. - PubMed
-
- Tohda C, Nakanishi R, Kadowaki M. Learning deficits and agenesis of synapses and myelinated axons in phosphoinositide-3 kinase-deficient mice. Neurosignals. 2006-2007;15(6):293–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
