

The depletion of F₁ subunit ε in yeast leads to an uncoupled respiratory phenotype that is rescued by mutations in the proton-translocating subunits of F₀
- PMID: 24451261
- PMCID: PMC3952849
- DOI: 10.1091/mbc.E13-02-0112
The depletion of F₁ subunit ε in yeast leads to an uncoupled respiratory phenotype that is rescued by mutations in the proton-translocating subunits of F₀
Abstract
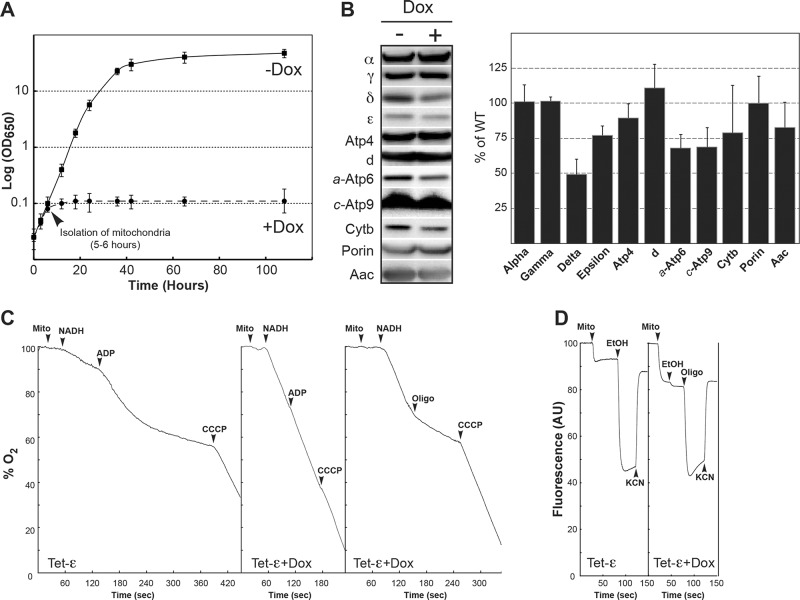
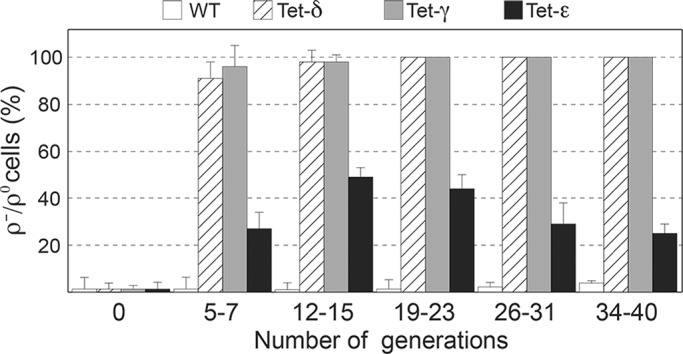
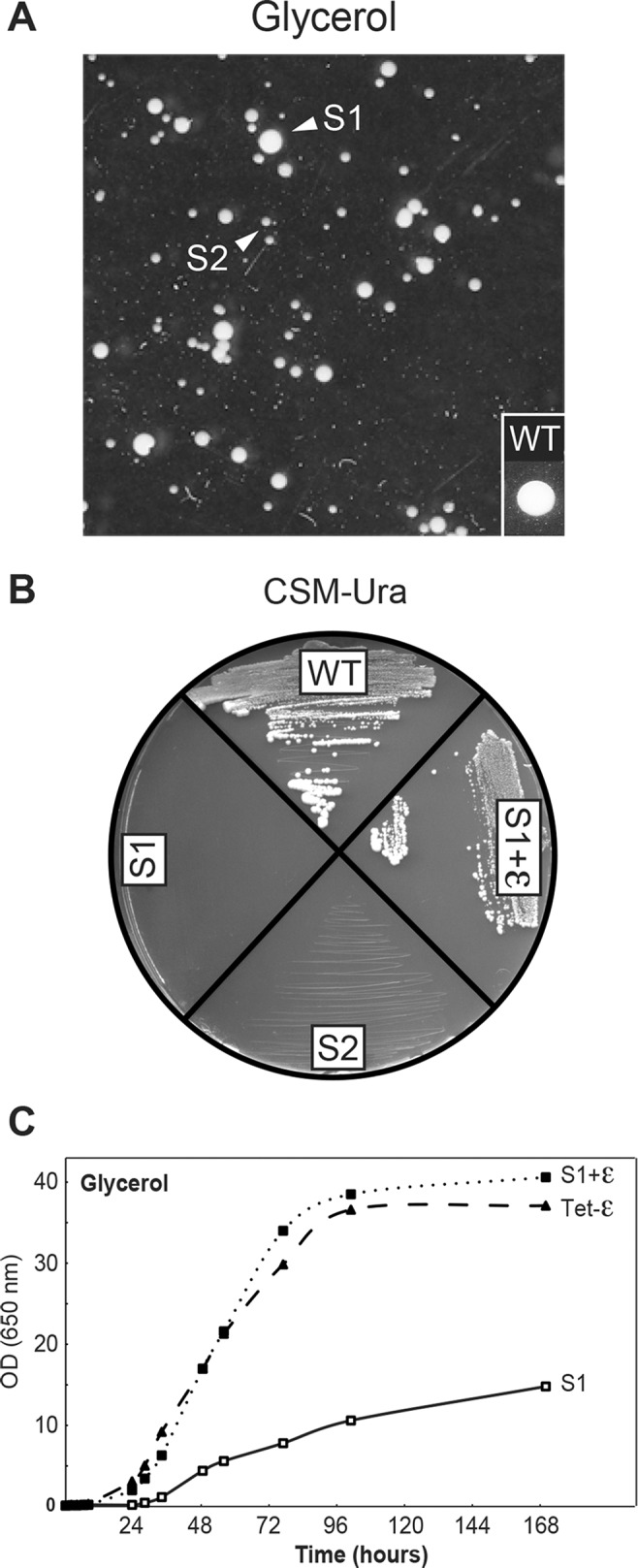
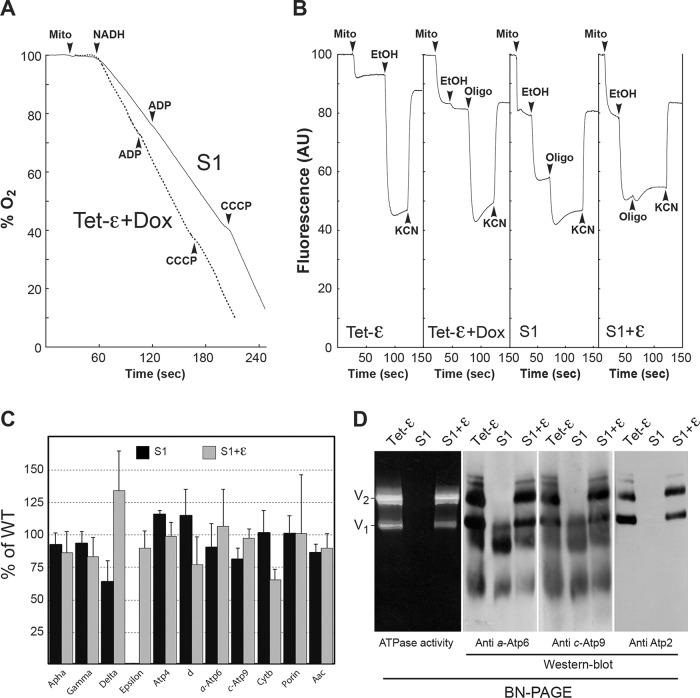
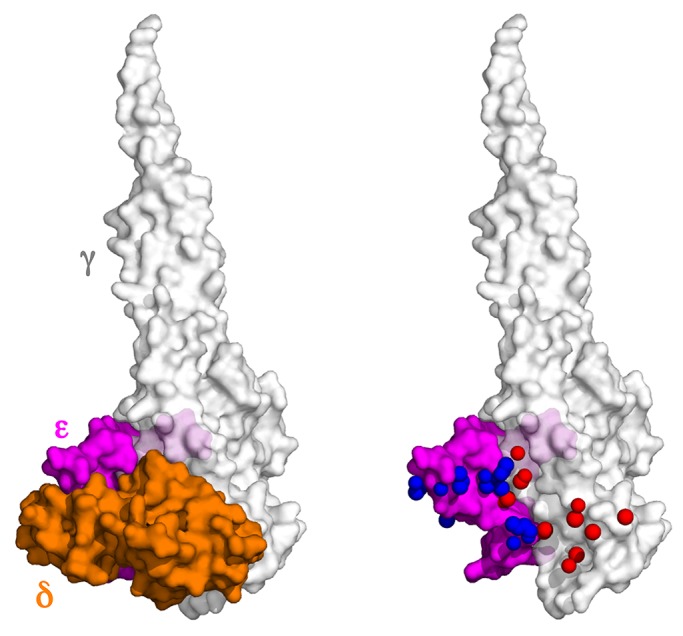
The central stalk of the ATP synthase is an elongated hetero-oligomeric structure providing a physical connection between the catalytic sites in F₁ and the proton translocation channel in F₀ for energy transduction between the two subdomains. The shape of the central stalk and relevance to energy coupling are essentially the same in ATP synthases from all forms of life, yet the protein composition of this domain changed during evolution of the mitochondrial enzyme from a two- to a three-subunit structure (γ, δ, ε). Whereas the mitochondrial γ- and δ-subunits are homologues of the bacterial central stalk proteins, the deliberate addition of subunit ε is poorly understood. Here we report that down-regulation of the gene (ATP15) encoding the ε-subunit rapidly leads to lethal F₀-mediated proton leaks through the membrane because of the loss of stability of the ATP synthase. The ε-subunit is thus essential for oxidative phosphorylation. Moreover, mutations in F₀ subunits a and c, which slow the proton translocation rate, are identified that prevent ε-deficient ATP synthases from dissipating the electrochemical potential. Cumulatively our data lead us to propose that the ε-subunit evolved to permit operation of the central stalk under the torque imposed at the normal speed of proton movement through mitochondrial F₀.
Figures
Similar articles
-
Manipulations in the peripheral stalk of the Saccharomyces cerevisiae F1F0-ATP synthase.J Biol Chem. 2011 Mar 25;286(12):10155-62. doi: 10.1074/jbc.M110.213447. Epub 2011 Jan 21. J Biol Chem. 2011. PMID: 21257750 Free PMC article.
-
ATP Synthase Subunit a Supports Permeability Transition in Yeast Lacking Dimerization Subunits and Modulates yPTP Conductance.Cell Physiol Biochem. 2020 Feb 27;54(2):211-229. doi: 10.33594/000000215. Cell Physiol Biochem. 2020. PMID: 32100973
-
Failure to assemble the alpha 3 beta 3 subcomplex of the ATP synthase leads to accumulation of the alpha and beta subunits within inclusion bodies and the loss of mitochondrial cristae in Saccharomyces cerevisiae.J Biol Chem. 2005 May 6;280(18):18386-92. doi: 10.1074/jbc.M410789200. Epub 2005 Feb 16. J Biol Chem. 2005. PMID: 15716275
-
Structures and interactions of proteins involved in the coupling function of the protonmotive F(o)F(1)-ATP synthase.Curr Protein Pept Sci. 2002 Aug;3(4):451-60. doi: 10.2174/1389203023380558. Curr Protein Pept Sci. 2002. PMID: 12370007 Review.
-
ATP synthase from Escherichia coli: Mechanism of rotational catalysis, and inhibition with the ε subunit and phytopolyphenols.Biochim Biophys Acta. 2016 Feb;1857(2):129-140. doi: 10.1016/j.bbabio.2015.11.005. Epub 2015 Nov 14. Biochim Biophys Acta. 2016. PMID: 26589785 Review.
Cited by
-
Understanding structure, function, and mutations in the mitochondrial ATP synthase.Microb Cell. 2015 Apr 1;2(4):105-125. doi: 10.15698/mic2015.04.197. Microb Cell. 2015. PMID: 25938092 Free PMC article.
-
Bedaquiline inhibits the yeast and human mitochondrial ATP synthases.Commun Biol. 2020 Aug 19;3(1):452. doi: 10.1038/s42003-020-01173-z. Commun Biol. 2020. PMID: 32814813 Free PMC article.
-
The ATP Synthase Deficiency in Human Diseases.Life (Basel). 2021 Apr 8;11(4):325. doi: 10.3390/life11040325. Life (Basel). 2021. PMID: 33917760 Free PMC article. Review.
-
Molecular basis of diseases induced by the mitochondrial DNA mutation m.9032T>C.Hum Mol Genet. 2023 Apr 6;32(8):1313-1323. doi: 10.1093/hmg/ddac292. Hum Mol Genet. 2023. PMID: 36434790 Free PMC article.
-
Biochemical investigation of a human pathogenic mutation in the nuclear ATP5E gene using yeast as a model.Front Genet. 2015 Apr 23;6:159. doi: 10.3389/fgene.2015.00159. eCollection 2015. Front Genet. 2015. PMID: 25954304 Free PMC article.
References
-
- Arselin G, Vaillier J, Graves PV, Velours J. ATP synthase of yeast mitochondria. Isolation of the subunit h and disruption of the ATP14 gene. J Biol Chem. 1996;271:20284–20290. - PubMed
-
- Devenish RJ, Prescott M, Rodgers AJ. The structure and function of mitochondrial F1F0-ATP synthases. Int Rev Cell Mol Biol. 2008;267:1–58. - PubMed
-
- Duvezin-Caubet S, Rak M, Lefebvre-Legendre L, Tetaud E, Bonnefoy N, di Rago JP. A “petite obligate” mutant of Saccharomyces cerevisiae: functional mtDNA is lethal in cells lacking the delta subunit of mitochondrial F1-ATPase. J Biol Chem. 2006;281:16305–16313. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
