

Histone deacetylase 2 (HDAC2) protein-dependent deacetylation of mortality factor 4-like 1 (MORF4L1) protein enhances its homodimerization
- PMID: 24451372
- PMCID: PMC3945369
- DOI: 10.1074/jbc.M113.527507
Histone deacetylase 2 (HDAC2) protein-dependent deacetylation of mortality factor 4-like 1 (MORF4L1) protein enhances its homodimerization
Erratum in
- J Biol Chem. 2014 Sep 5;289(36):24943
Abstract
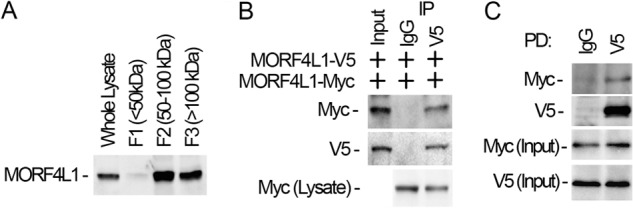
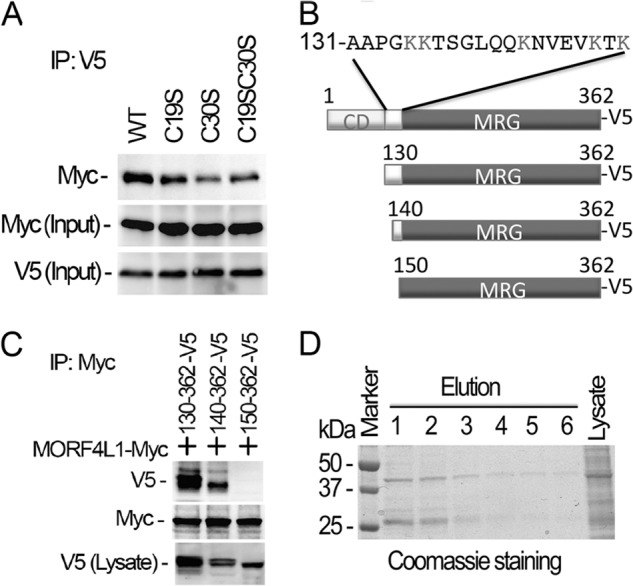
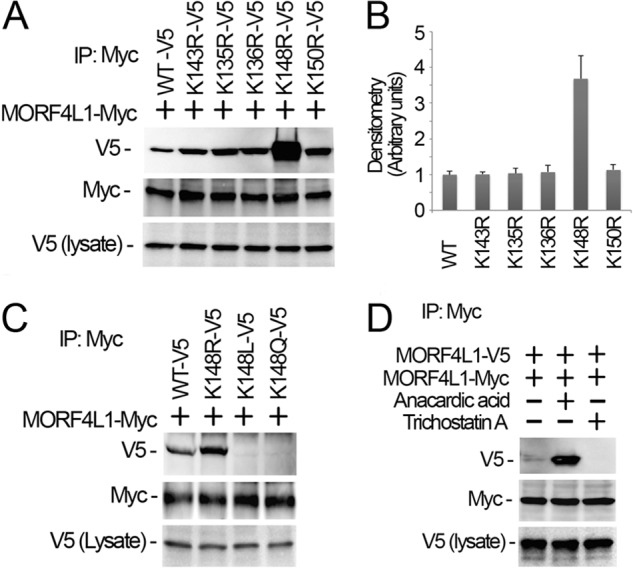
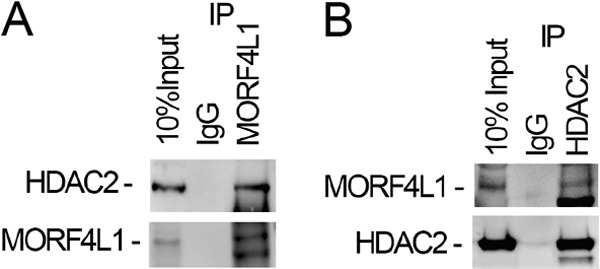
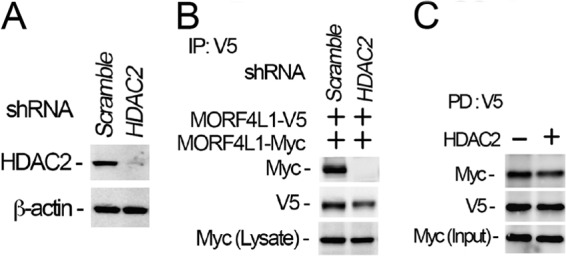
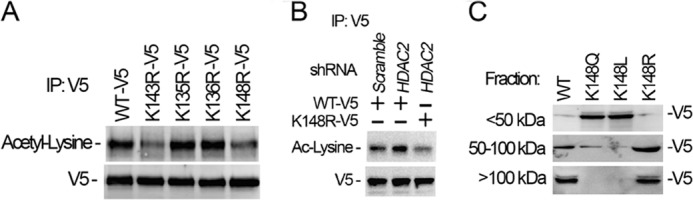
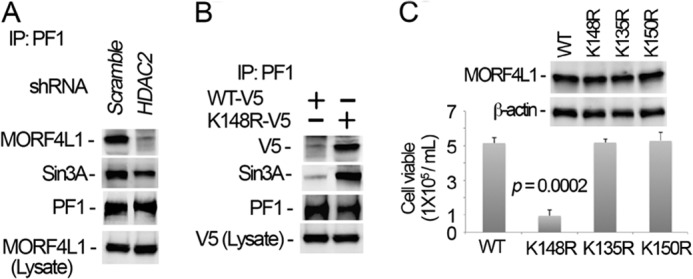
Histone acetyltransferase mortality factor 4-like 1 (MORF4L1) is a relatively new histone acetyltransferase component that exists as a homodimer to exert its epigenetic function. The mechanism of MORF4L1 self-assembly is unknown. Here we report that Lys-148 deacetylation is indispensable for facilitating MORF4L1 self-assembly into a homodimeric unit. Among a stretch of ∼10 amino acids in the NH2 terminus between the chromodomain and MORF4-related gene (MRG) domain within MORF4L1, Lys-148 is normally acetylated. Substitution of Lys-148 with arginine augments MORF4L1 self-assembly. However, acetylation mimics of MORF4L1, including K148L and K148Q, abolished its self-assembly of the histone acetyltransferase component. HDAC2, a deacetylase, interacts with and keeps MORF4L1 in a deacetylation status at Lys(148) that triggers MORF4L1 self-assembly. Knockdown of HDAC2 reduces MORF4L1 self-assembly. HDAC2-dependent deacetylation of MORF4L1 enhances MORF4L1 homodimerization, thus facilitating the functionality of complex formation to repress cell proliferation.
Keywords: Epigenetics; Histone Acetylase; Histone Deacetylase; Posttranslational Modification; Proliferation; Protein Complexes; Protein Structure; Site-directed Mutagenesis.
Figures
Similar articles
-
Regulation of transcription factor YY1 by acetylation and deacetylation.Mol Cell Biol. 2001 Sep;21(17):5979-91. doi: 10.1128/MCB.21.17.5979-5991.2001. Mol Cell Biol. 2001. PMID: 11486036 Free PMC article.
-
Histone deacetylase 2 (HDAC2) regulates chromosome segregation and kinetochore function via H4K16 deacetylation during oocyte maturation in mouse.PLoS Genet. 2013;9(3):e1003377. doi: 10.1371/journal.pgen.1003377. Epub 2013 Mar 14. PLoS Genet. 2013. PMID: 23516383 Free PMC article.
-
Trans-regulation of histone deacetylase activities through acetylation.J Biol Chem. 2009 Dec 11;284(50):34901-10. doi: 10.1074/jbc.M109.038356. Epub 2009 Oct 11. J Biol Chem. 2009. PMID: 19822520 Free PMC article.
-
HDAC1 and HDAC2 in mouse oocytes and preimplantation embryos: Specificity versus compensation.Cell Death Differ. 2016 Jul;23(7):1119-27. doi: 10.1038/cdd.2016.31. Epub 2016 Apr 15. Cell Death Differ. 2016. PMID: 27082454 Free PMC article. Review.
-
Histone deacetylase-2 and airway disease.Ther Adv Respir Dis. 2009 Oct;3(5):235-43. doi: 10.1177/1753465809348648. Epub 2009 Oct 7. Ther Adv Respir Dis. 2009. PMID: 19812111 Review.
Cited by
-
MORF4L1 suppresses cell proliferation, migration and invasion by increasing p21 and E-cadherin expression in nasopharyngeal carcinoma.Oncol Lett. 2019 Jan;17(1):294-302. doi: 10.3892/ol.2018.9588. Epub 2018 Oct 16. Oncol Lett. 2019. PMID: 30655767 Free PMC article.
-
Oncogenic Gene CNOT7 Promotes Progression and Induces Poor Prognosis of Glioma.Mol Biotechnol. 2025 Jul;67(7):2607-2616. doi: 10.1007/s12033-024-01223-5. Epub 2024 Jul 10. Mol Biotechnol. 2025. PMID: 38985240 Free PMC article.
-
Systems Biology Approaches to Investigate Genetic and Epigenetic Molecular Progression Mechanisms for Identifying Gene Expression Signatures in Papillary Thyroid Cancer.Int J Mol Sci. 2019 May 23;20(10):2536. doi: 10.3390/ijms20102536. Int J Mol Sci. 2019. PMID: 31126066 Free PMC article.
-
Structural and functional insights into the epigenetic regulator MRG15.Acta Pharmacol Sin. 2024 May;45(5):879-889. doi: 10.1038/s41401-023-01211-6. Epub 2024 Jan 8. Acta Pharmacol Sin. 2024. PMID: 38191914 Free PMC article. Review.
-
Molecular Mechanisms of Vascular Damage During Lung Injury.Adv Exp Med Biol. 2021;1304:95-107. doi: 10.1007/978-3-030-68748-9_6. Adv Exp Med Biol. 2021. PMID: 34019265 Free PMC article. Review.
References
-
- Qi H. H., Sarkissian M., Hu G. Q., Wang Z., Bhattacharjee A., Gordon D. B., Gonzales M., Lan F., Ongusaha P. P., Huarte M., Yaghi N. K., Lim H., Garcia B. A., Brizuela L., Zhao K., Roberts T. M., Shi Y. (2010) Histone H4K20/H3K9 demethylase PHF8 regulates zebrafish brain and craniofacial development. Nature 466, 503–507 - PMC - PubMed
-
- Bertram M. J., Pereira-Smith O. M. (2001) Conservation of the MORF4 related gene family. Identification of a new chromo domain subfamily and novel protein motif. Gene 266, 111–121 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
