

Induction of virulence gene expression in Staphylococcus aureus by pulmonary surfactant
- PMID: 24452679
- PMCID: PMC3993393
- DOI: 10.1128/IAI.01635-13
Induction of virulence gene expression in Staphylococcus aureus by pulmonary surfactant
Abstract
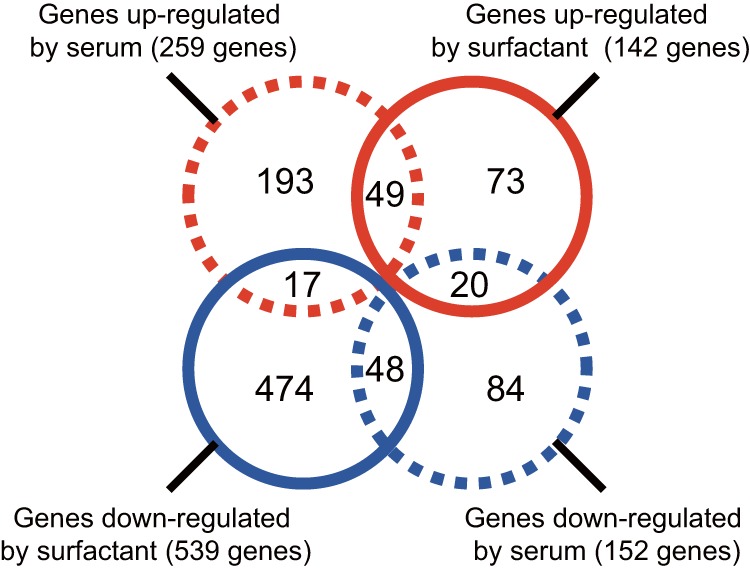
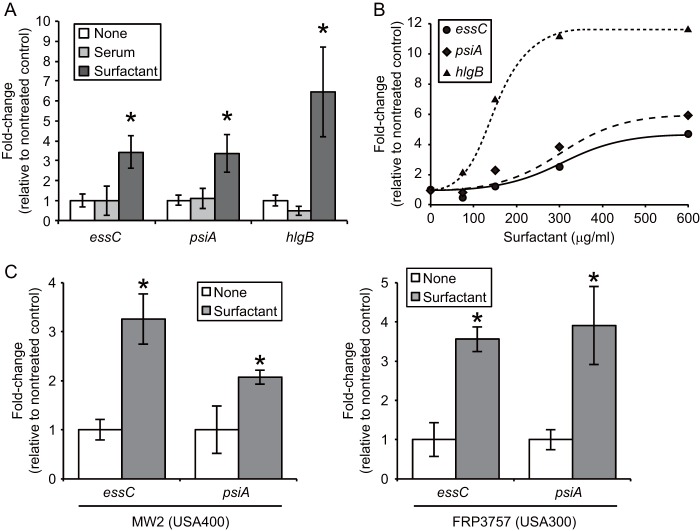
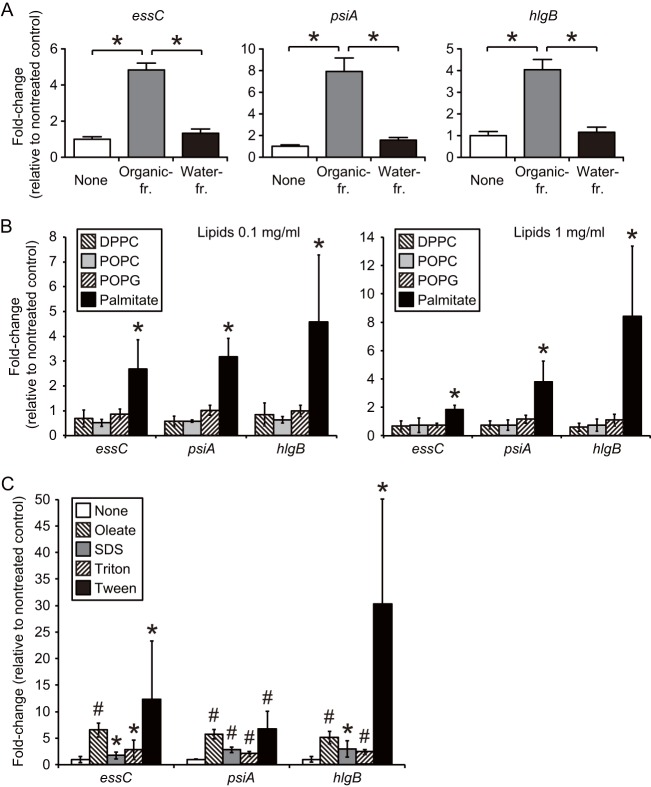
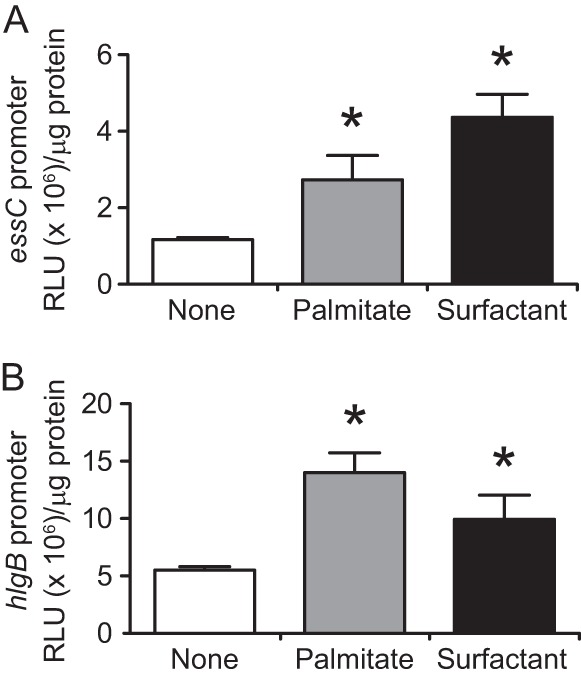
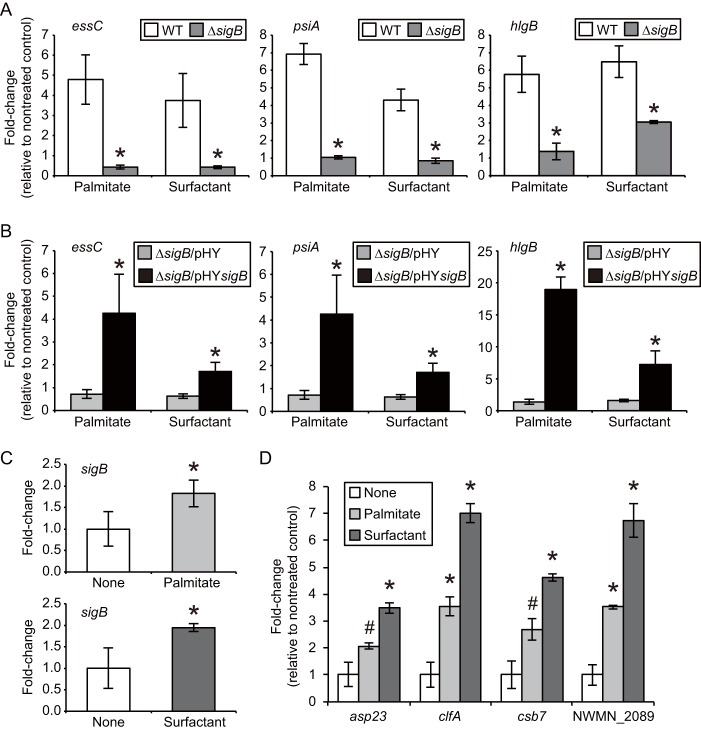
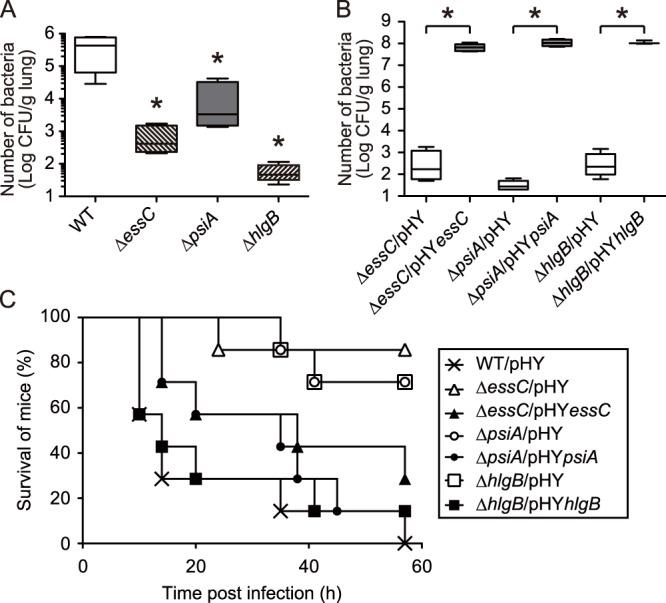
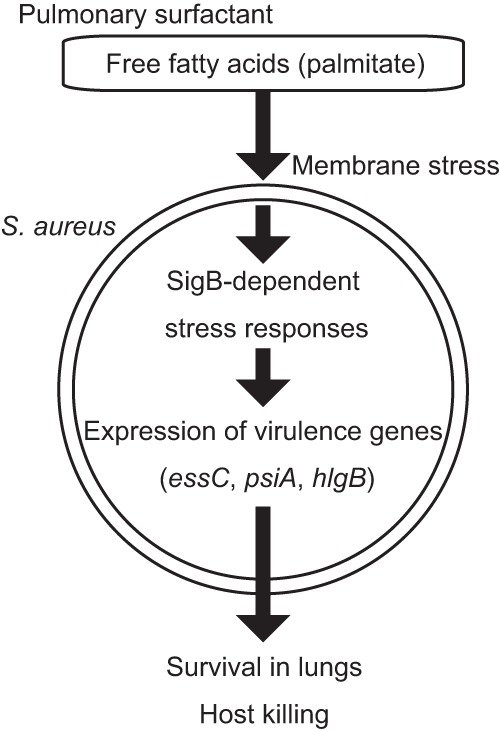
We performed a genomewide analysis using a next-generation sequencer to investigate the effect of pulmonary surfactant on gene expression in Staphylococcus aureus, a clinically important opportunistic pathogen. RNA sequence (RNA-seq) analysis of bacterial transcripts at late log phase revealed 142 genes that were upregulated >2-fold following the addition of pulmonary surfactant to the culture medium. Among these genes, we confirmed by quantitative reverse transcription-PCR analysis that mRNA amounts for genes encoding ESAT-6 secretion system C (EssC), an unknown hypothetical protein (NWMN_0246; also called pulmonary surfactant-inducible factor A [PsiA] in this study), and hemolysin gamma subunit B (HlgB) were increased 3- to 10-fold by the surfactant treatment. Among the major constituents of pulmonary surfactant, i.e., phospholipids and palmitate, only palmitate, which is the most abundant fatty acid in the pulmonary surfactant and a known antibacterial substance, stimulated the expression of these three genes. Moreover, these genes were also induced by supplementing the culture with detergents. The induction of gene expression by surfactant or palmitate was not observed in a disruption mutant of the sigB gene, which encodes an alternative sigma factor involved in bacterial stress responses. Furthermore, each disruption mutant of the essC, psiA, and hlgB genes showed attenuation of both survival in the lung and host-killing ability in a murine pneumonia model. These findings suggest that S. aureus resists membrane stress caused by free fatty acids present in the pulmonary surfactant through the regulation of virulence gene expression, which contributes to its pathogenesis within the lungs of the host animal.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
