

A ribozyme that triphosphorylates RNA 5'-hydroxyl groups
- PMID: 24452796
- PMCID: PMC3985629
- DOI: 10.1093/nar/gkt1405
A ribozyme that triphosphorylates RNA 5'-hydroxyl groups
Abstract
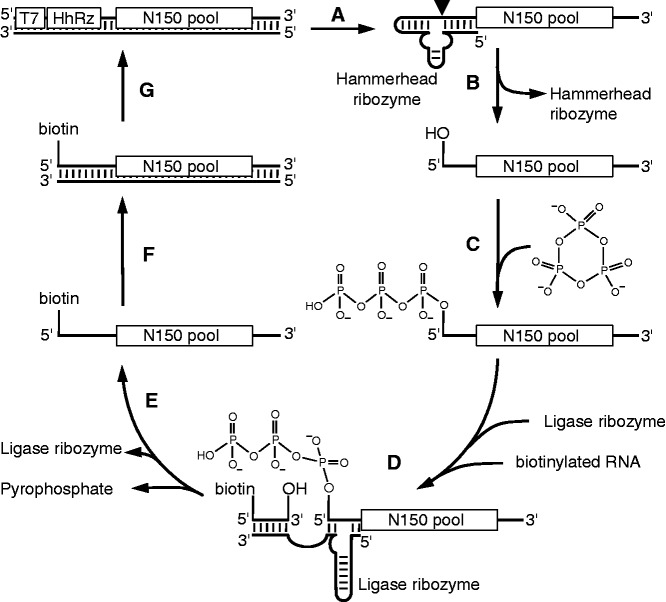
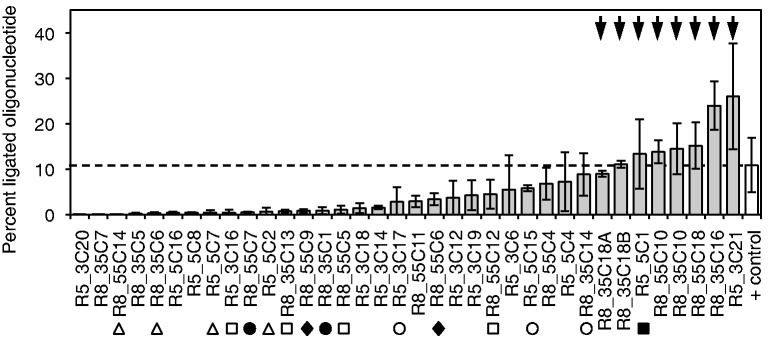
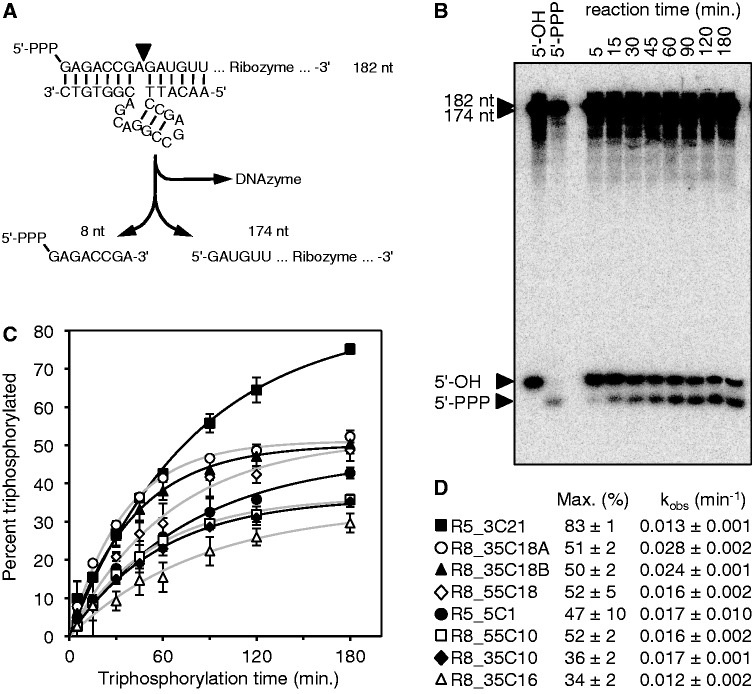
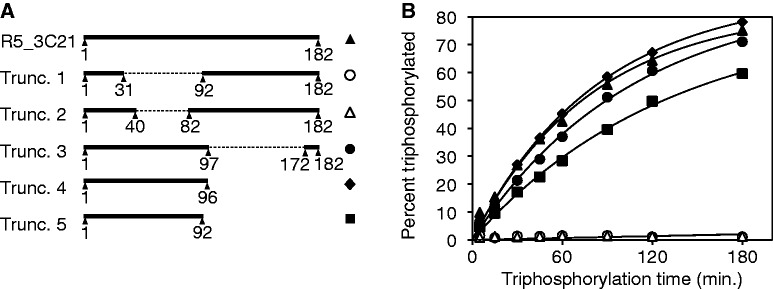
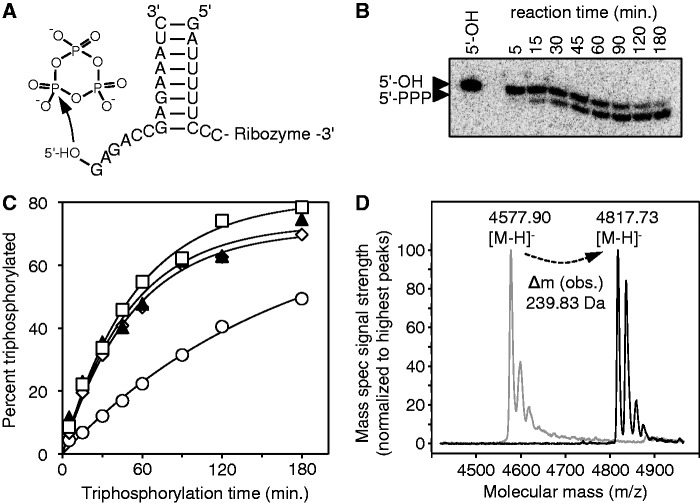
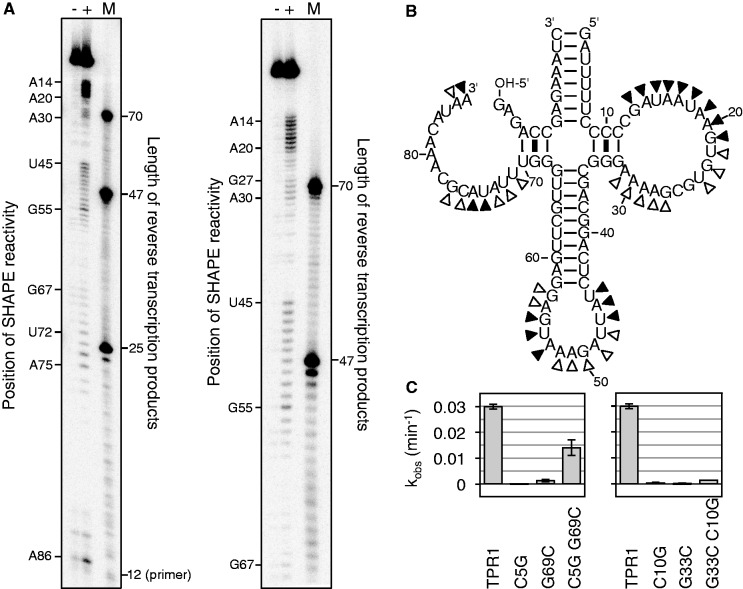
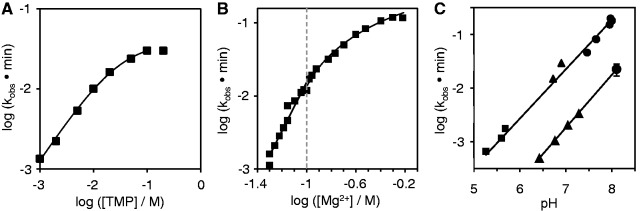
The RNA world hypothesis describes a stage in the early evolution of life in which RNA served as genome and as the only genome-encoded catalyst. To test whether RNA world organisms could have used cyclic trimetaphosphate as an energy source, we developed an in vitro selection strategy for isolating ribozymes that catalyze the triphosphorylation of RNA 5'-hydroxyl groups with trimetaphosphate. Several active sequences were isolated, and one ribozyme was analyzed in more detail. The ribozyme was truncated to 96 nt, while retaining full activity. It was converted to a trans-format and reacted with rates of 0.16 min(-1) under optimal conditions. The secondary structure appears to contain a four-helical junction motif. This study showed that ribozymes can use trimetaphosphate to triphosphorylate RNA 5'-hydroxyl groups and suggested that RNA world organisms could have used trimetaphosphate as their energy source.
Figures
References
-
- Woese CR. The Genetic Code. The Molecular Basis for Genetic Expression. New York: Harper & Row; 1967.
-
- Crick FHC. The origin of the genetic code. J. Mol. Biol. 1968;38:367–379. - PubMed
-
- Orgel LE. Evolution of the genetic apparatus. J. Mol. Biol. 1968;38:381–393. - PubMed
-
- Szostak JW. The eightfold path to non-enzymatic RNA replication. J. Syst. Chem. 2012;3:2.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
