

Expanding the zinc-finger recombinase repertoire: directed evolution and mutational analysis of serine recombinase specificity determinants
- PMID: 24452803
- PMCID: PMC3985619
- DOI: 10.1093/nar/gkt1389
Expanding the zinc-finger recombinase repertoire: directed evolution and mutational analysis of serine recombinase specificity determinants
Abstract
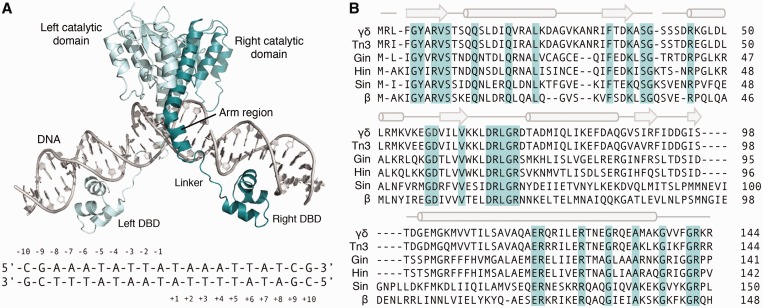
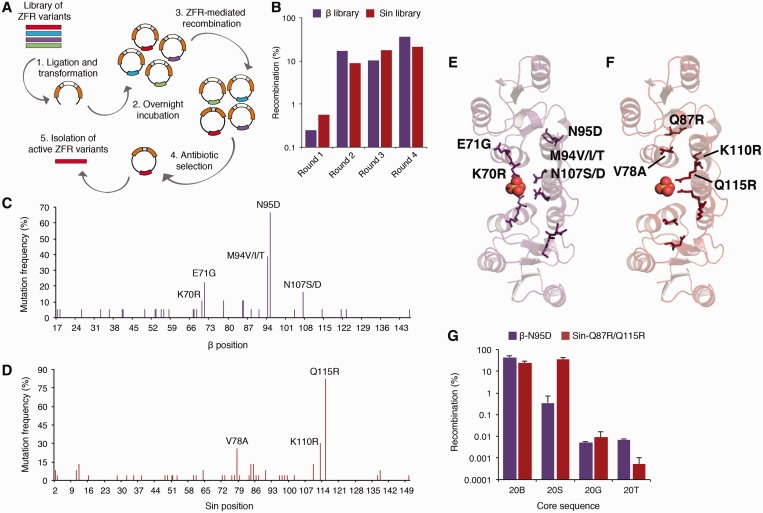
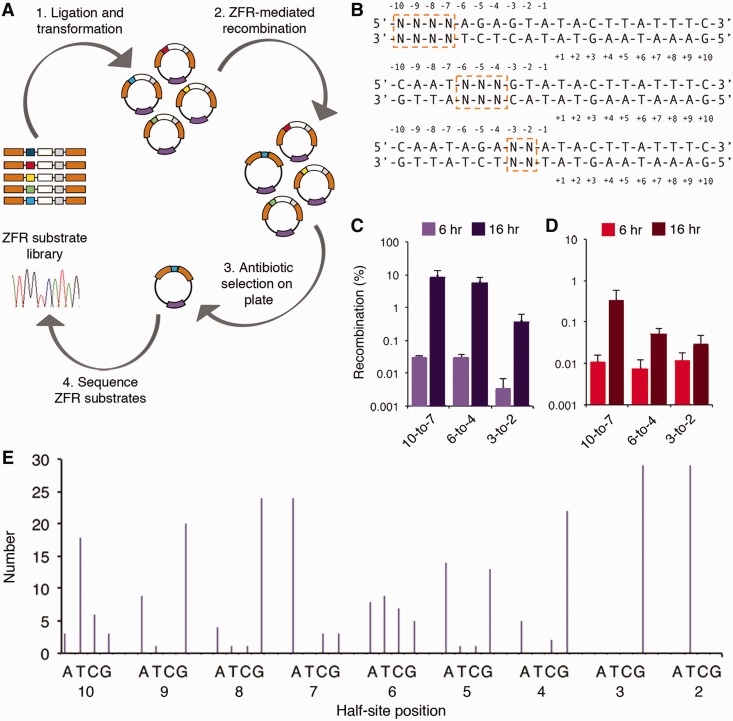
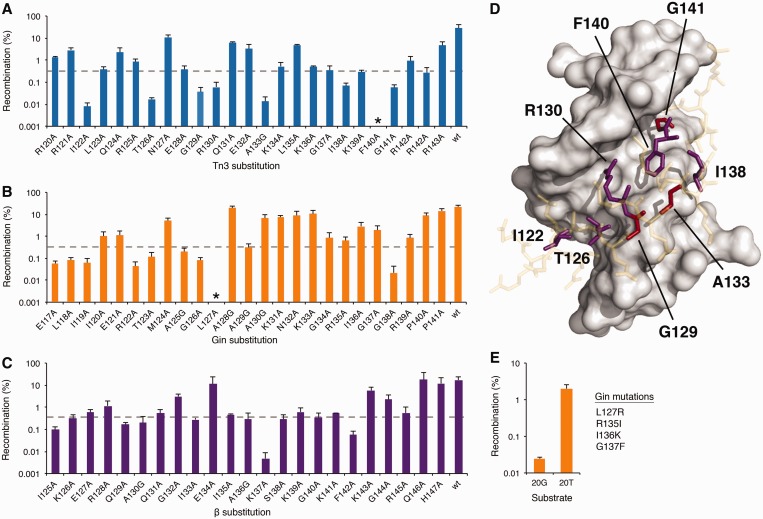
The serine recombinases are a diverse family of modular enzymes that promote high-fidelity DNA rearrangements between specific target sites. Replacement of their native DNA-binding domains with custom-designed Cys₂-His₂ zinc-finger proteins results in the creation of engineered zinc-finger recombinases (ZFRs) capable of achieving targeted genetic modifications. The flexibility afforded by zinc-finger domains enables the design of hybrid recombinases that recognize a wide variety of potential target sites; however, this technology remains constrained by the strict recognition specificities imposed by the ZFR catalytic domains. In particular, the ability to fully reprogram serine recombinase catalytic specificity has been impeded by conserved base requirements within each recombinase target site and an incomplete understanding of the factors governing DNA recognition. Here we describe an approach to complement the targeting capacity of ZFRs. Using directed evolution, we isolated mutants of the β and Sin recombinases that specifically recognize target sites previously outside the scope of ZFRs. Additionally, we developed a genetic screen to determine the specific base requirements for site-specific recombination and showed that specificity profiling enables the discovery of unique genomic ZFR substrates. Finally, we conducted an extensive and family-wide mutational analysis of the serine recombinase DNA-binding arm region and uncovered a diverse network of residues that confer target specificity. These results demonstrate that the ZFR repertoire is extensible and highlights the potential of ZFRs as a class of flexible tools for targeted genome engineering.
Figures
References
-
- Bibikova M, Beumer K, Trautman JK, Carroll D. Enhancing gene targeting with designed zinc finger nucleases. Science. 2003;300:764. - PubMed
-
- Miller JC, Tan S, Qiao G, Barlow KA, Wang J, Xia DF, Meng X, Paschon DE, Leung E, Hinkley SJ, et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011;29:143–148. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
