

Phosphoenolpyruvate carboxylase identified as a key enzyme in erythrocytic Plasmodium falciparum carbon metabolism
- PMID: 24453970
- PMCID: PMC3894211
- DOI: 10.1371/journal.ppat.1003876
Phosphoenolpyruvate carboxylase identified as a key enzyme in erythrocytic Plasmodium falciparum carbon metabolism
Abstract
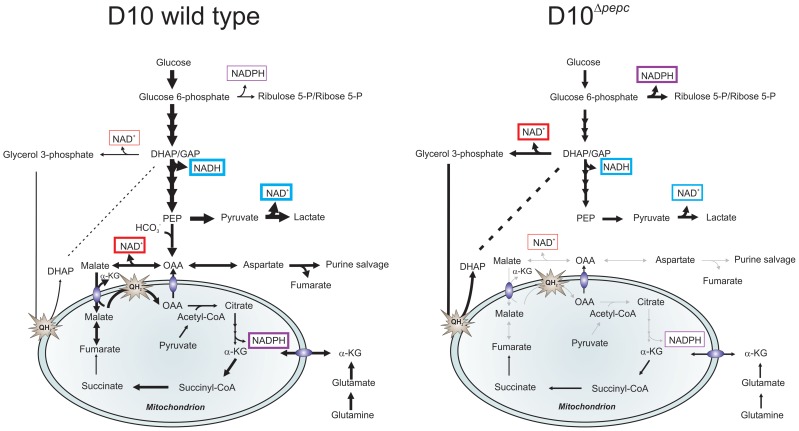
Phospoenolpyruvate carboxylase (PEPC) is absent from humans but encoded in the Plasmodium falciparum genome, suggesting that PEPC has a parasite-specific function. To investigate its importance in P. falciparum, we generated a pepc null mutant (D10(Δpepc) ), which was only achievable when malate, a reduction product of oxaloacetate, was added to the growth medium. D10(Δpepc) had a severe growth defect in vitro, which was partially reversed by addition of malate or fumarate, suggesting that pepc may be essential in vivo. Targeted metabolomics using (13)C-U-D-glucose and (13)C-bicarbonate showed that the conversion of glycolytically-derived PEP into malate, fumarate, aspartate and citrate was abolished in D10(Δpepc) and that pentose phosphate pathway metabolites and glycerol 3-phosphate were present at increased levels. In contrast, metabolism of the carbon skeleton of (13)C,(15)N-U-glutamine was similar in both parasite lines, although the flux was lower in D10(Δpepc); it also confirmed the operation of a complete forward TCA cycle in the wild type parasite. Overall, these data confirm the CO2 fixing activity of PEPC and suggest that it provides metabolites essential for TCA cycle anaplerosis and the maintenance of cytosolic and mitochondrial redox balance. Moreover, these findings imply that PEPC may be an exploitable target for future drug discovery.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
Regulation of the supply of oxaloacetate for mitochondrial metabolism via phosphoenolpyruvate carboxylase in barley leaf protoplasts. II. Effects of metabolites on PEPC activity at different activation states of the protein.Biochim Biophys Acta. 1996 Apr 17;1289(3):351-61. doi: 10.1016/0304-4165(95)00165-4. Biochim Biophys Acta. 1996. PMID: 8620019
-
Genetic investigation of tricarboxylic acid metabolism during the Plasmodium falciparum life cycle.Cell Rep. 2015 Apr 7;11(1):164-74. doi: 10.1016/j.celrep.2015.03.011. Epub 2015 Apr 2. Cell Rep. 2015. PMID: 25843709 Free PMC article.
-
Regulation of the supply of cytosolic oxaloacetate for mitochondrial metabolism via phosphoenolpyruvate carboxylase in barley leaf protoplasts. I. The effect of covalent modification on PEPC activity, pH response, and kinetic properties.Biochim Biophys Acta. 1996 Apr 17;1289(3):343-50. doi: 10.1016/0304-4165(95)00164-6. Biochim Biophys Acta. 1996. PMID: 8620018
-
The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs.Biochem J. 2011 May 15;436(1):15-34. doi: 10.1042/BJ20110078. Biochem J. 2011. PMID: 21524275 Review.
-
[Functions of plant phosphoenolpyruvate carboxylase and its applications for genetic engineering].Sheng Wu Gong Cheng Xue Bao. 2011 Dec;27(12):1702-10. Sheng Wu Gong Cheng Xue Bao. 2011. PMID: 22506410 Review. Chinese.
Cited by
-
Metabolic changes accompanying the loss of fumarate hydratase and malate-quinone oxidoreductase in the asexual blood stage of Plasmodium falciparum.J Biol Chem. 2022 May;298(5):101897. doi: 10.1016/j.jbc.2022.101897. Epub 2022 Apr 6. J Biol Chem. 2022. PMID: 35398098 Free PMC article.
-
Purinergic Receptor Antagonists Inhibit Hemolysis Induced by Clostridium perfringens Alpha Toxin.Pathogens. 2024 May 27;13(6):454. doi: 10.3390/pathogens13060454. Pathogens. 2024. PMID: 38921752 Free PMC article.
-
Plasmodium falciparum LipB mutants display altered redox and carbon metabolism in asexual stages and cannot complete sporogony in Anopheles mosquitoes.Int J Parasitol. 2021 May;51(6):441-453. doi: 10.1016/j.ijpara.2020.10.011. Epub 2021 Mar 11. Int J Parasitol. 2021. PMID: 33713652 Free PMC article.
-
β-Carboline Derivatives Tackling Malaria: Biological Evaluation and Docking Analysis.ACS Omega. 2020 Jul 13;5(29):17993-18006. doi: 10.1021/acsomega.0c01256. eCollection 2020 Jul 28. ACS Omega. 2020. PMID: 32743172 Free PMC article.
-
Metabolic alterations in the erythrocyte during blood-stage development of the malaria parasite.Malar J. 2020 Feb 27;19(1):94. doi: 10.1186/s12936-020-03174-z. Malar J. 2020. PMID: 32103749 Free PMC article.
References
-
- WHO, (2012) World Malaria Report 2012, World Health Organisation, Geneva, Switzerland, 59.
-
- Foth BJ, Stimmler LM, Handman E, Crabb BS, Hodder AN, et al. (2005) The malaria parasite Plasmodium falciparum has only one pyruvate dehydrogenase complex, which is located in the apicoplast. Mol Microbiol 55: 39–53. - PubMed
-
- Pei Y, Tarun AS, Vaughan AM, Herman RW, Soliman JM, et al. (2010) Plasmodium pyruvate dehydrogenase activity is only essential for the parasite's progression from liver infection to blood infection. Mol Microbiol 75: 957–971. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
