

Aerodynamic characteristics of a feathered dinosaur measured using physical models. Effects of form on static stability and control effectiveness
- PMID: 24454820
- PMCID: PMC3893193
- DOI: 10.1371/journal.pone.0085203
Aerodynamic characteristics of a feathered dinosaur measured using physical models. Effects of form on static stability and control effectiveness
Abstract
We report the effects of posture and morphology on the static aerodynamic stability and control effectiveness of physical models based on the feathered dinosaur, [Formula: see text]Microraptor gui, from the Cretaceous of China. Postures had similar lift and drag coefficients and were broadly similar when simplified metrics of gliding were considered, but they exhibited different stability characteristics depending on the position of the legs and the presence of feathers on the legs and the tail. Both stability and the function of appendages in generating maneuvering forces and torques changed as the glide angle or angle of attack were changed. These are significant because they represent an aerial environment that may have shifted during the evolution of directed aerial descent and other aerial behaviors. Certain movements were particularly effective (symmetric movements of the wings and tail in pitch, asymmetric wing movements, some tail movements). Other appendages altered their function from creating yaws at high angle of attack to rolls at low angle of attack, or reversed their function entirely. While [Formula: see text]M. gui lived after [Formula: see text]Archaeopteryx and likely represents a side experiment with feathered morphology, the general patterns of stability and control effectiveness suggested from the manipulations of forelimb, hindlimb and tail morphology here may help understand the evolution of flight control aerodynamics in vertebrates. Though these results rest on a single specimen, as further fossils with different morphologies are tested, the findings here could be applied in a phylogenetic context to reveal biomechanical constraints on extinct flyers arising from the need to maneuver.
Conflict of interest statement
Figures
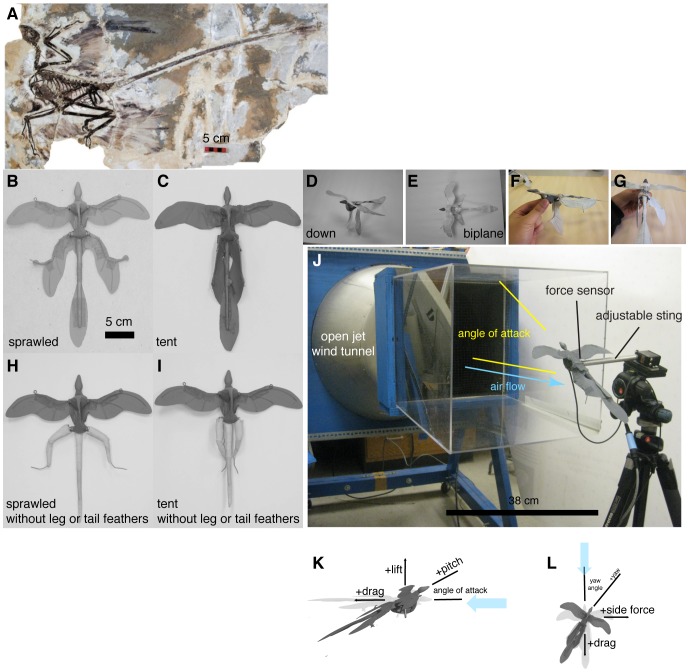
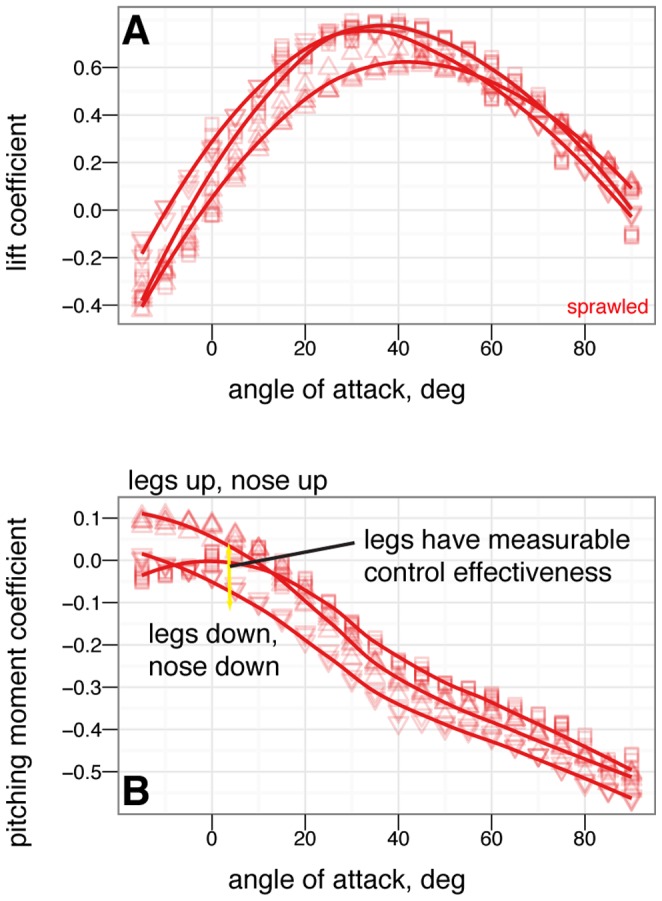
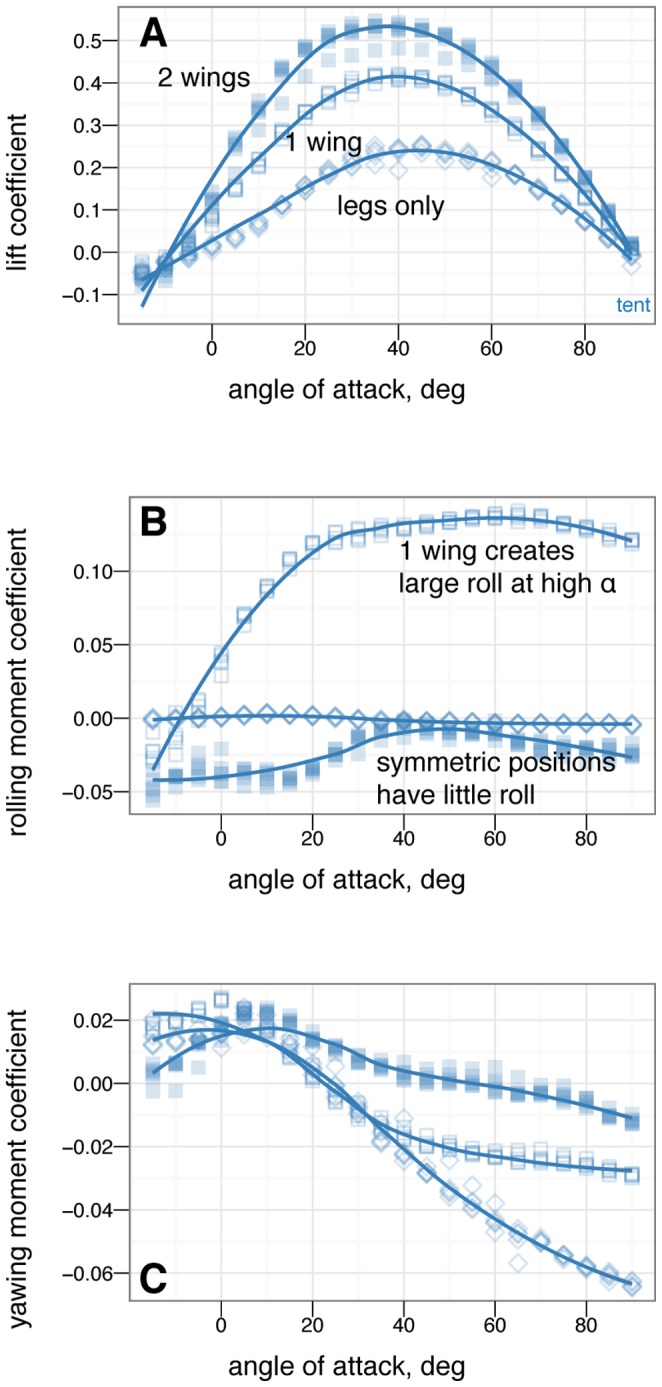
 M. gui, scale model wingspan 20 cm, snout-vent-length 8 cm. Reconstruction postures, B-I, used for constructing physical models: B, sprawled, after ; C, tent, after , ; D, legs-down, after ; E, biplane, after . F-I additional manipulations: F, asymmetric leg posture with 9090
M. gui, scale model wingspan 20 cm, snout-vent-length 8 cm. Reconstruction postures, B-I, used for constructing physical models: B, sprawled, after ; C, tent, after , ; D, legs-down, after ; E, biplane, after . F-I additional manipulations: F, asymmetric leg posture with 9090 leg mismatch ( arabesque ); G, example asymmetric leg posture with 45
leg mismatch ( arabesque ); G, example asymmetric leg posture with 45 dihedral on one leg ( dégagé ), H, sprawled without leg or tail feathers; I, tent without leg or tail feathers. J, test setup; K, sign conventions, rotation angles, and definitions for model testing, after , , .
dihedral on one leg ( dégagé ), H, sprawled without leg or tail feathers; I, tent without leg or tail feathers. J, test setup; K, sign conventions, rotation angles, and definitions for model testing, after , , .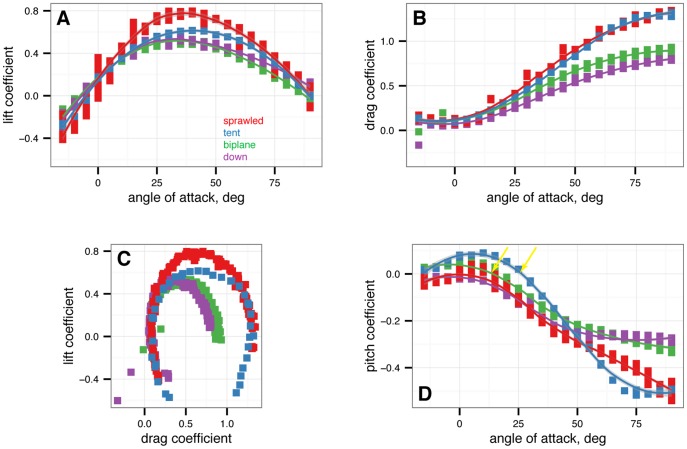
 from –15
from –15 to 90
to 90 in 5
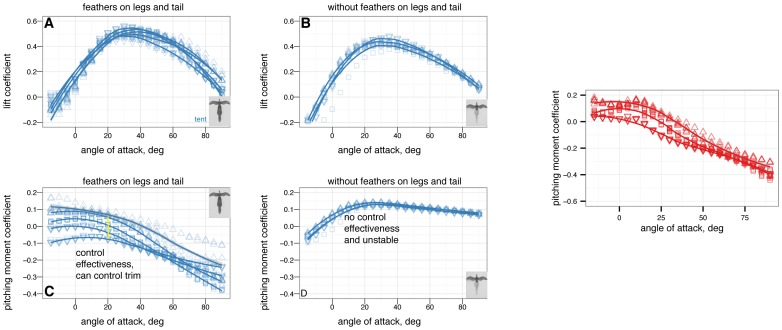
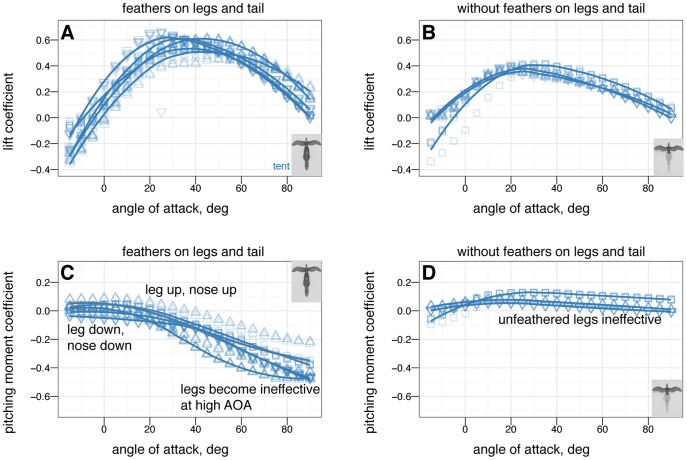
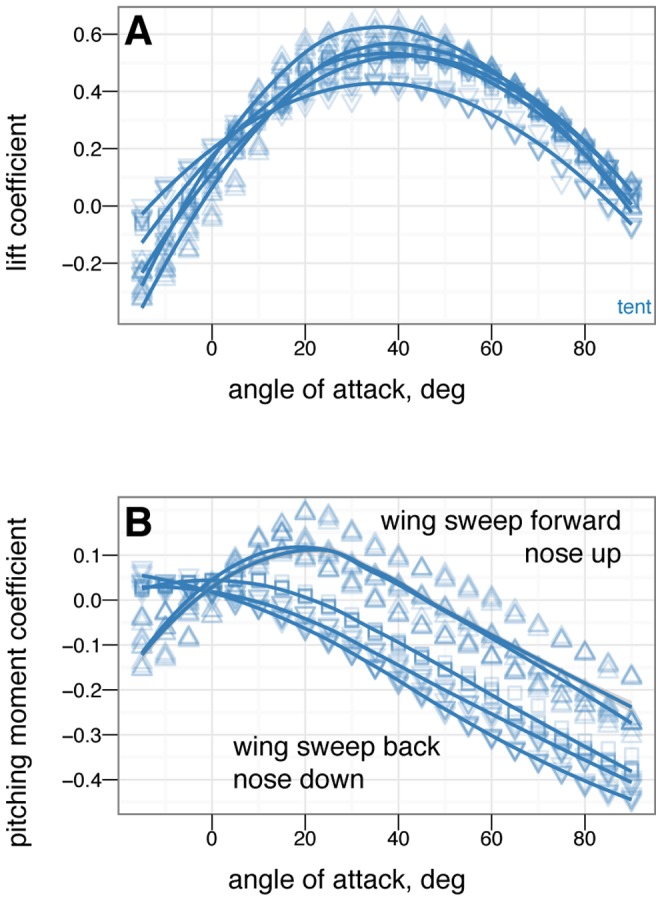
in 5 increments, with five or more replicates per treatment. A, Lift coefficient. B, Drag coefficient. C, Lift drag polars. D, Pitching moment coefficient. Stable angles of attack, which cross
increments, with five or more replicates per treatment. A, Lift coefficient. B, Drag coefficient. C, Lift drag polars. D, Pitching moment coefficient. Stable angles of attack, which cross  with negative slope, for tent (blue) and biplane (green) postures identified with yellow arrows.
with negative slope, for tent (blue) and biplane (green) postures identified with yellow arrows.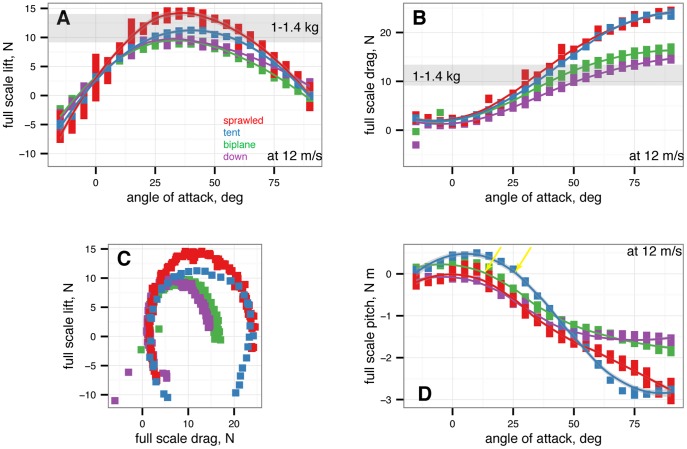
 from –15
from –15 to 90
to 90 in 5
in 5 increments, with five or more replicates per treatment. Gray band indicates weight range of
increments, with five or more replicates per treatment. Gray band indicates weight range of  M. gui. A, Full scale lift at 12
M. gui. A, Full scale lift at 12  , all models. B, Full scale drag at 12
, all models. B, Full scale drag at 12  , all models. C, Lift-drag polars. D, Full scale pitching moment at 12
, all models. C, Lift-drag polars. D, Full scale pitching moment at 12  versus angle of attack, all models. Stable angles of attack for tent (blue) and biplane (green) indicated.
versus angle of attack, all models. Stable angles of attack for tent (blue) and biplane (green) indicated.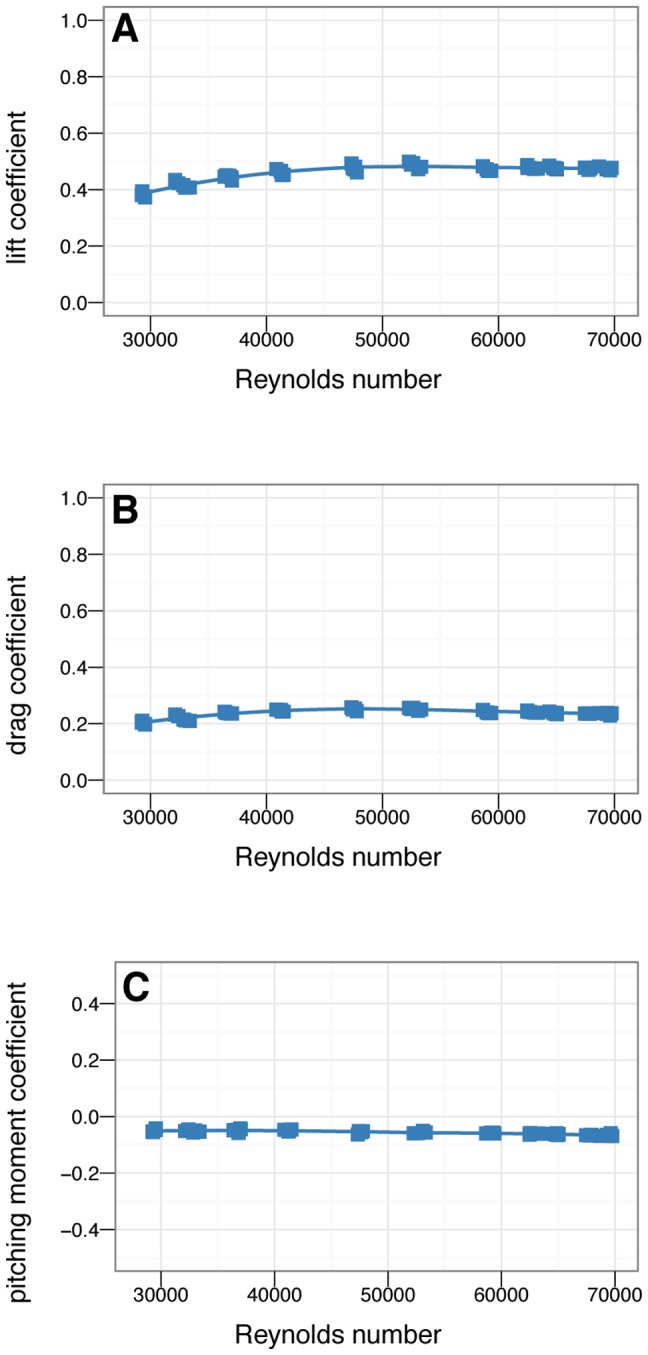
 Archaeopteryx. Moment coefficients are constant over the range shown.
Archaeopteryx. Moment coefficients are constant over the range shown.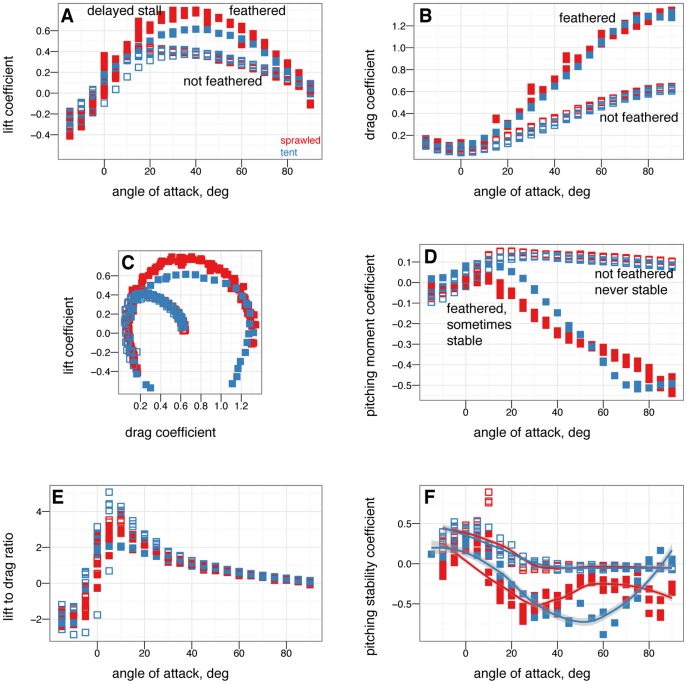
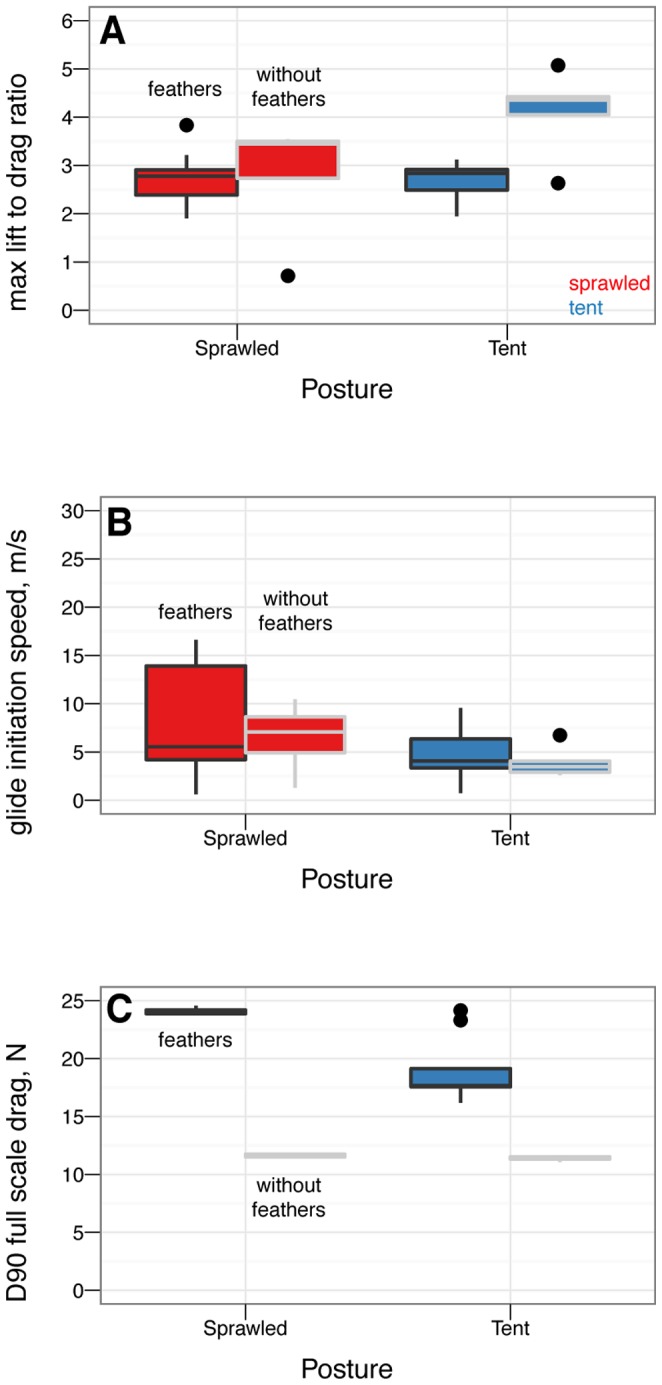
 ), however, this improvement is never achieved because the tent posture is never stable without leg feathers. B, Minimum glide speed, by sprawled and tent postures with and without feathers. There are no differences in minimum glide speed between postures (ANOVA,
), however, this improvement is never achieved because the tent posture is never stable without leg feathers. B, Minimum glide speed, by sprawled and tent postures with and without feathers. There are no differences in minimum glide speed between postures (ANOVA,  ). C, Parachuting drag, by sprawled and tent postures with and without feathers. There are significant differences in parachuting drag between postures (ANOVA,
). C, Parachuting drag, by sprawled and tent postures with and without feathers. There are significant differences in parachuting drag between postures (ANOVA,  ), however, the straight-down parachuting position is not stable.
), however, the straight-down parachuting position is not stable.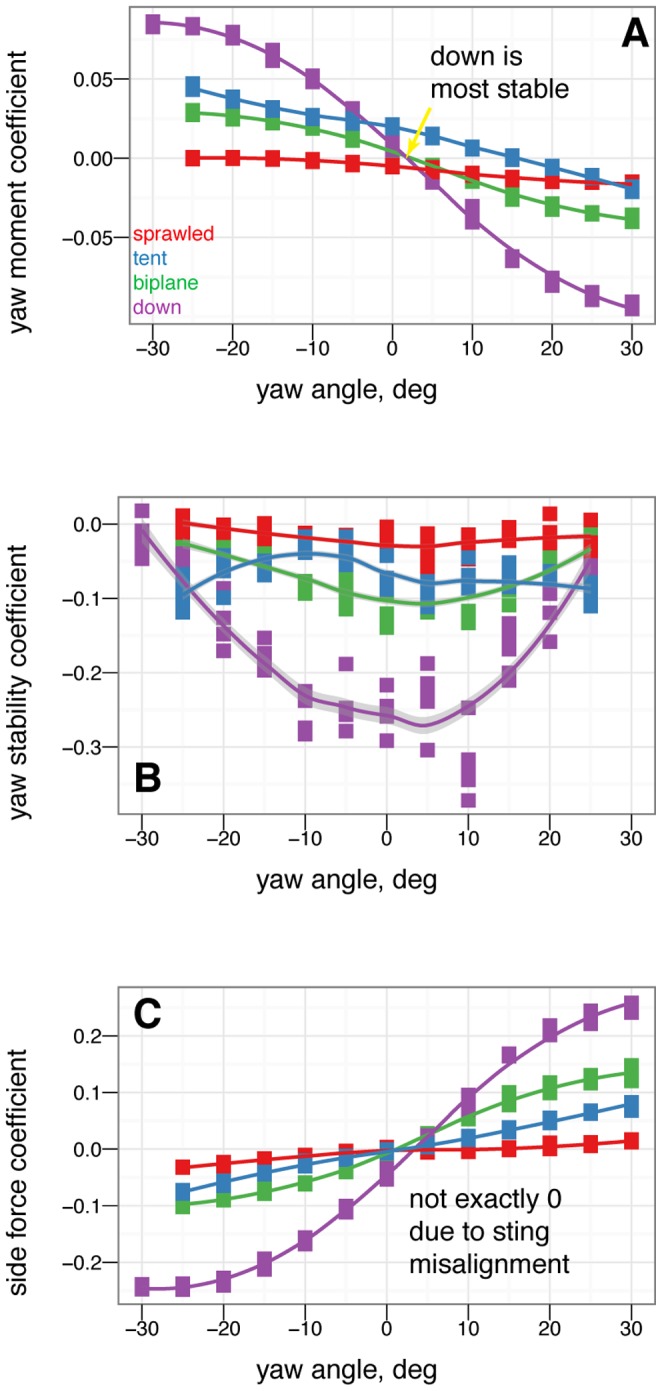
 ). Color represents the base posture: red for sprawled, blue for tent, green for biplane, and purple for down.
). Color represents the base posture: red for sprawled, blue for tent, green for biplane, and purple for down.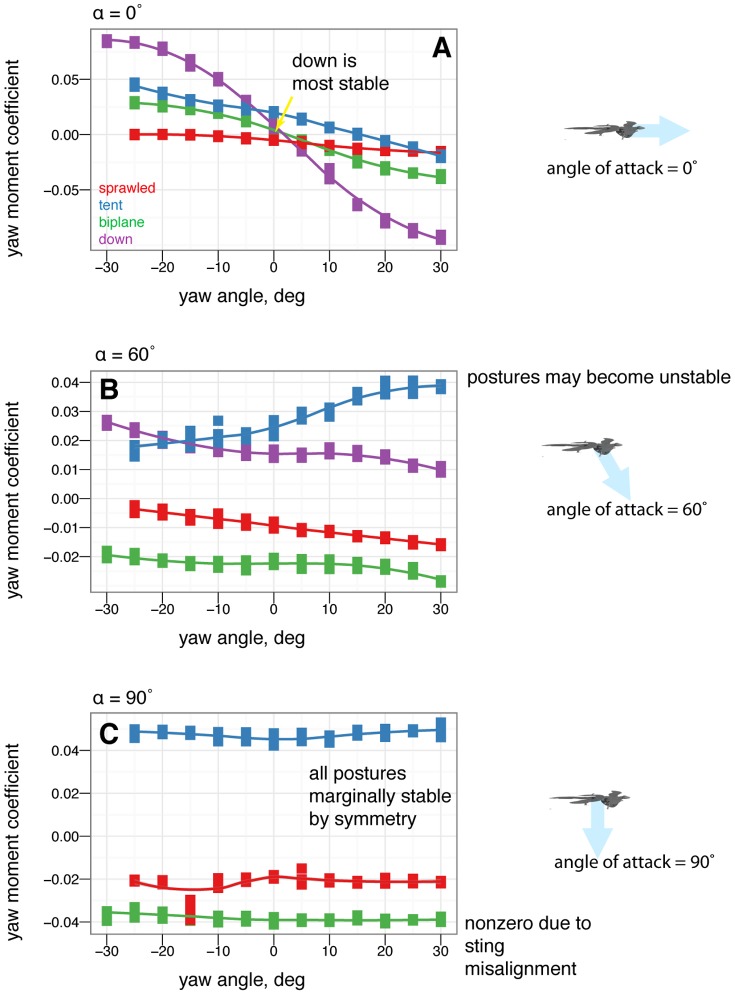
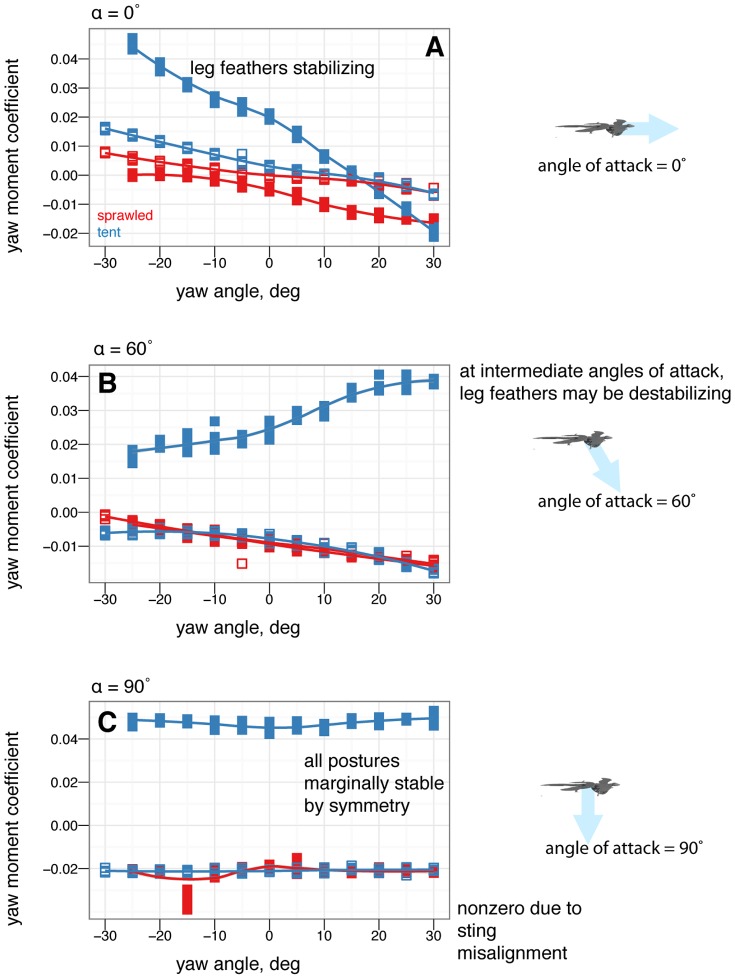
 , some postures are more stable in yaw than others. B, At 60
, some postures are more stable in yaw than others. B, At 60 , postures that were stable at 0
, postures that were stable at 0 may go unstable, such as tent posture. C, At 90
may go unstable, such as tent posture. C, At 90 , all postures are marginally stable due to symmetry (lines flat, yawing does not alter position relative to flow). Color represents the base posture: red for sprawled, blue for tent. Organisms may have navigated this transition from 90
, all postures are marginally stable due to symmetry (lines flat, yawing does not alter position relative to flow). Color represents the base posture: red for sprawled, blue for tent. Organisms may have navigated this transition from 90 to 0
to 0 .
.
 , some feathered-leg postures are more stable in yaw than others. B, At 60
, some feathered-leg postures are more stable in yaw than others. B, At 60 , postures that were stable at 0
, postures that were stable at 0 may go unstable, such as tent posture with leg feathers. C, At 90
may go unstable, such as tent posture with leg feathers. C, At 90 , all postures are marginally stable due to symmetry. Color represents the base posture: red for sprawled, blue for tent, green for biplane, and purple for down.
, all postures are marginally stable due to symmetry. Color represents the base posture: red for sprawled, blue for tent, green for biplane, and purple for down.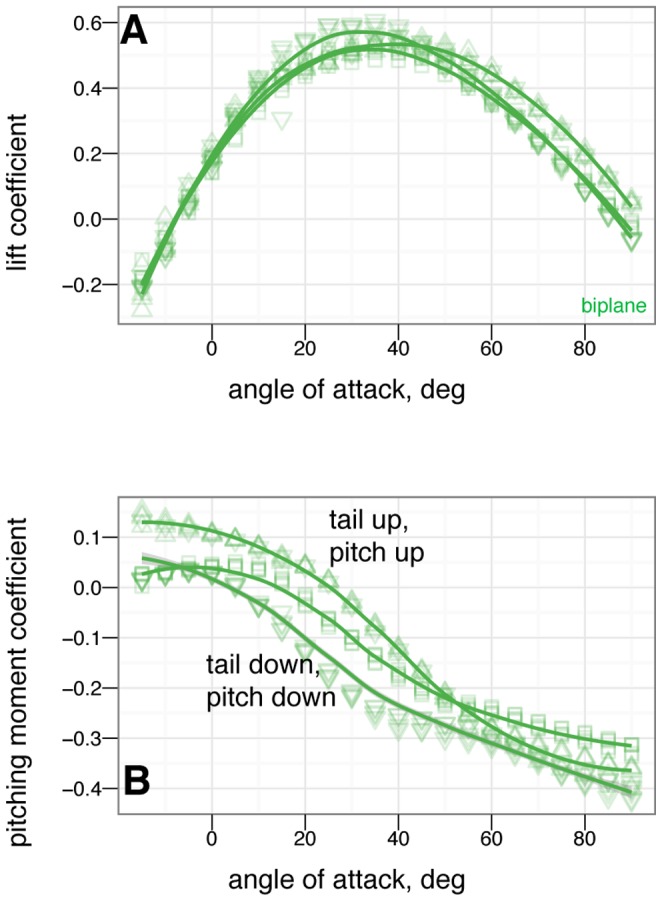
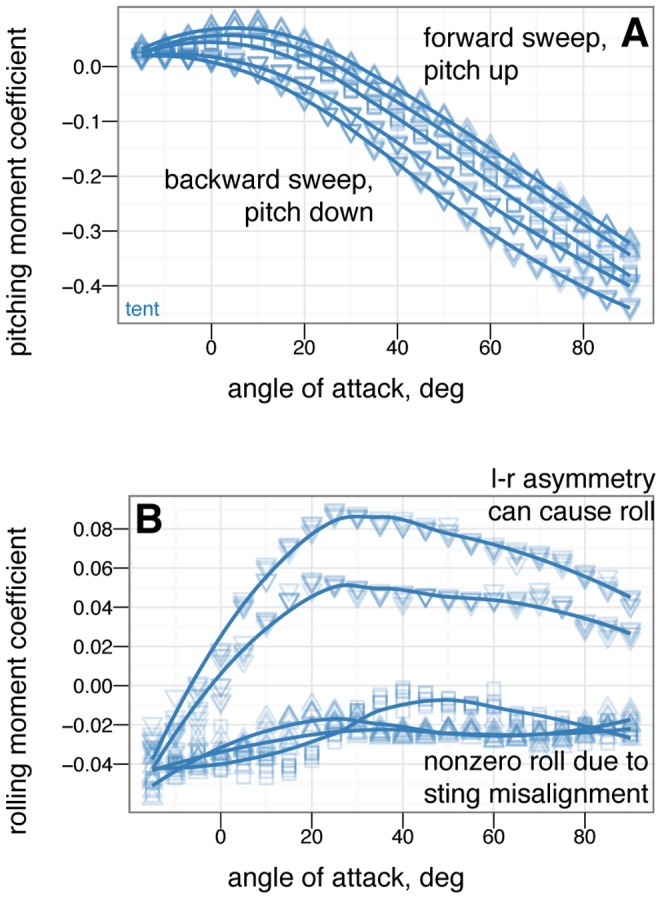
 . The small effect on lift suggests the tail is primarily effective because of moments generated by its long length.
. The small effect on lift suggests the tail is primarily effective because of moments generated by its long length.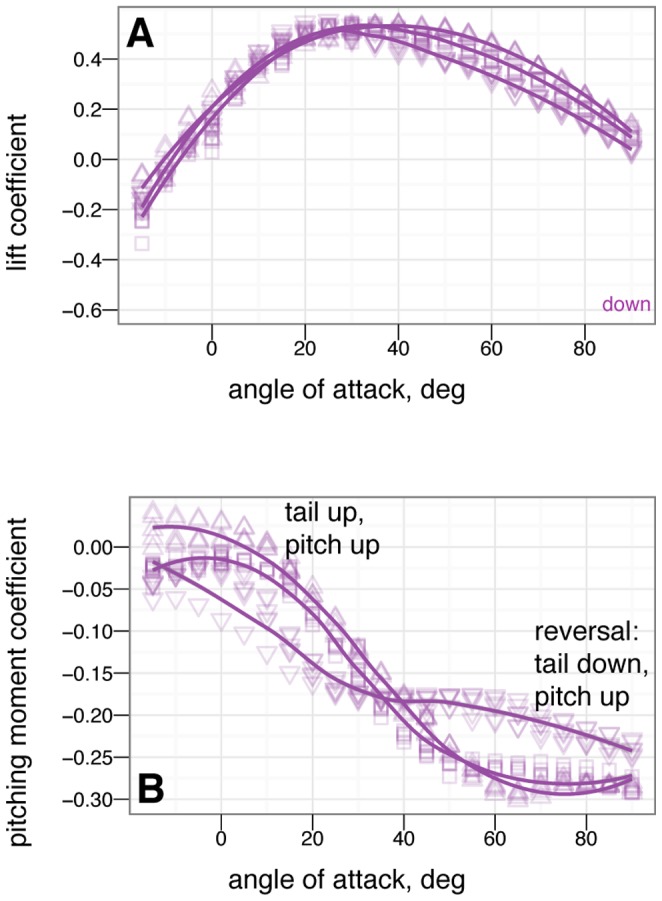
 tail movement. At high angle of attack, the tail experiences reversal in which tail down produces nose up moments / tail up produces nose down moments.
tail movement. At high angle of attack, the tail experiences reversal in which tail down produces nose up moments / tail up produces nose down moments.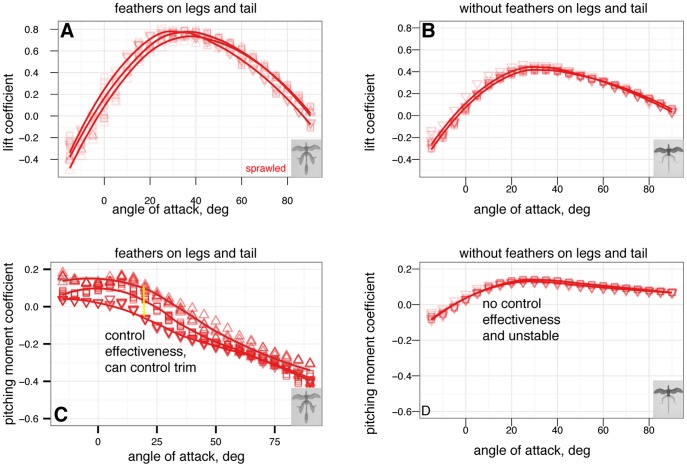
 angle of attack.
angle of attack.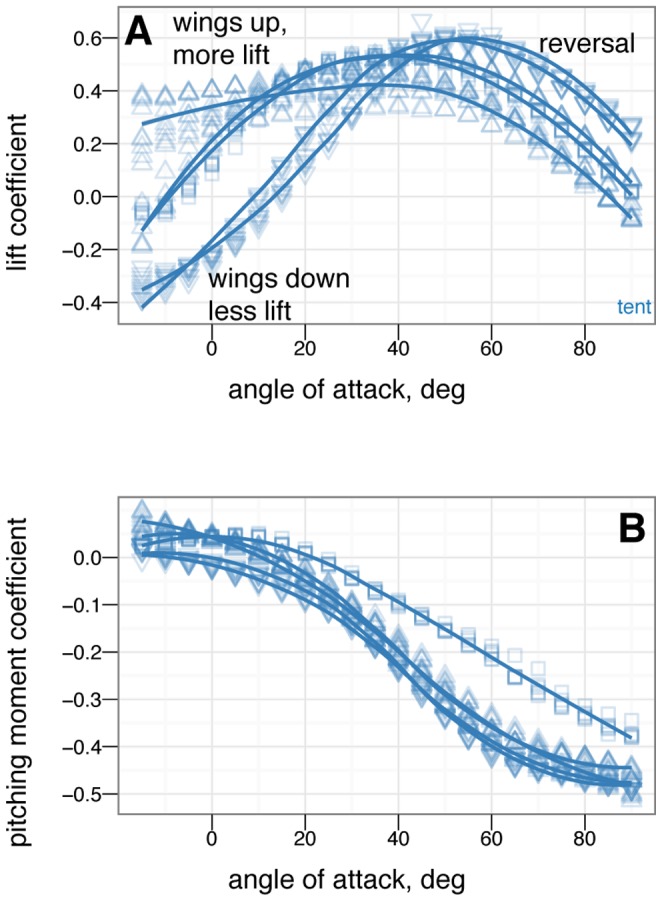
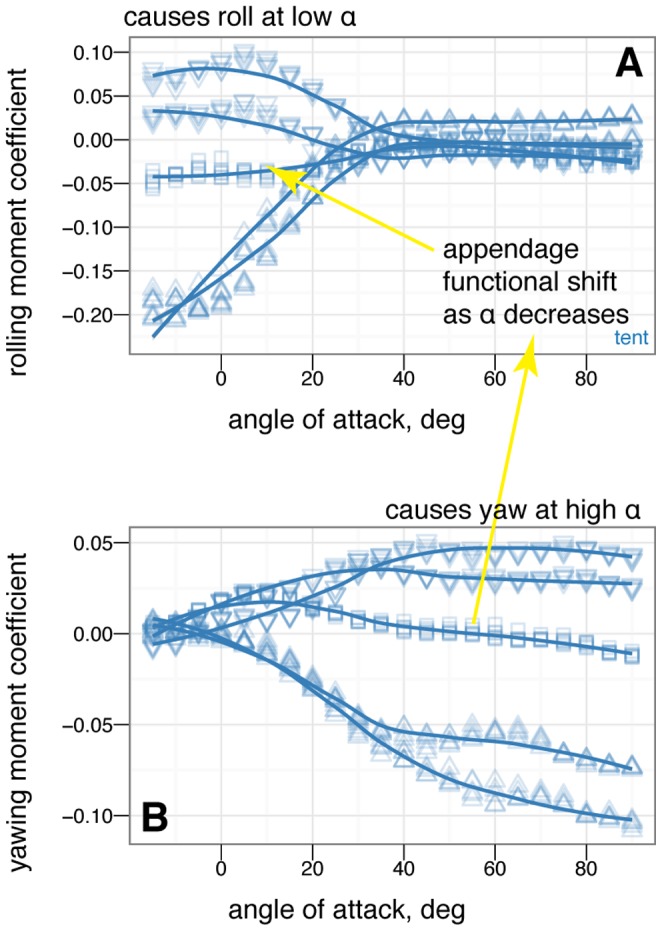
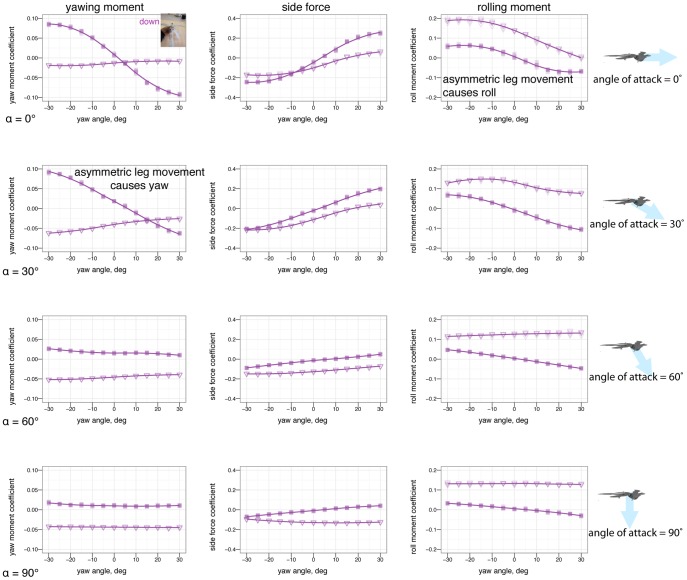
 dihedral (down arrow). Placing one leg at a dihedral is destabilizing in yaw and produces side force and rolling and yawing moments due to the asymmetry.
dihedral (down arrow). Placing one leg at a dihedral is destabilizing in yaw and produces side force and rolling and yawing moments due to the asymmetry.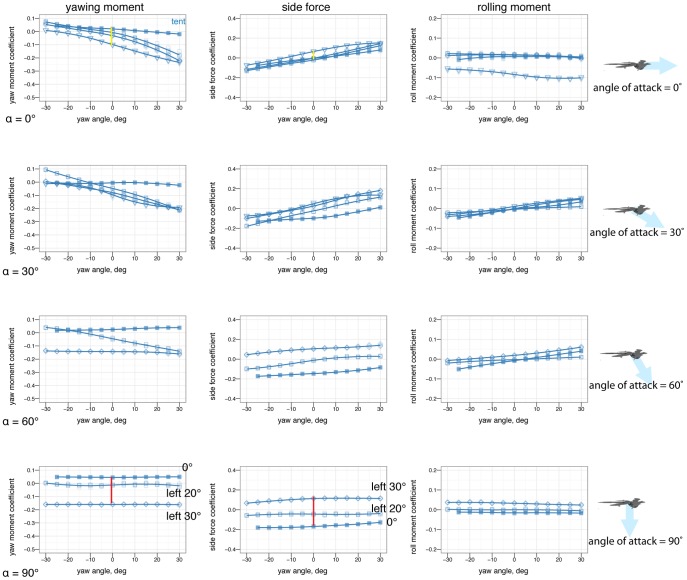
 left (open square), tail 20
left (open square), tail 20 left (open triangle), tail 30
left (open triangle), tail 30 left (open diamond). The tail is effective at creating yawing moments but at low angles of attack it is shadowed by the body and larger movements are needed (yellow versus red lines).
left (open diamond). The tail is effective at creating yawing moments but at low angles of attack it is shadowed by the body and larger movements are needed (yellow versus red lines).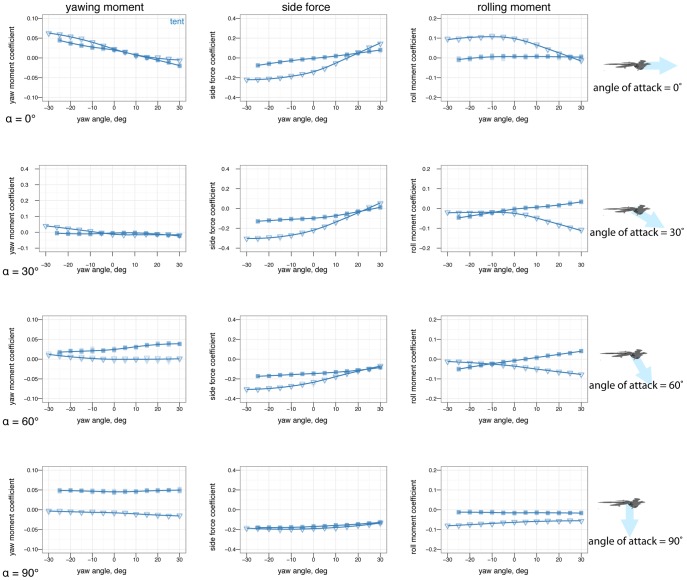
Similar articles
-
Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight.Nat Commun. 2013;4:2489. doi: 10.1038/ncomms3489. Nat Commun. 2013. PMID: 24048346
-
Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui.Proc Natl Acad Sci U S A. 2007 Jan 30;104(5):1576-80. doi: 10.1073/pnas.0609975104. Epub 2007 Jan 22. Proc Natl Acad Sci U S A. 2007. PMID: 17242354 Free PMC article.
-
Using physical models to study the gliding performance of extinct animals.Integr Comp Biol. 2011 Dec;51(6):1002-18. doi: 10.1093/icb/icr112. Epub 2011 Sep 21. Integr Comp Biol. 2011. PMID: 21937667
-
Morphology and distribution of scales, dermal ossifications, and other non-feather integumentary structures in non-avialan theropod dinosaurs.Biol Rev Camb Philos Soc. 2022 Jun;97(3):960-1004. doi: 10.1111/brv.12829. Epub 2022 Jan 6. Biol Rev Camb Philos Soc. 2022. PMID: 34991180 Review.
-
Rarity of molt evidence in early pennaraptoran dinosaurs suggests annual molt evolved later among Neornithes.Commun Biol. 2023 Jul 3;6(1):687. doi: 10.1038/s42003-023-05048-x. Commun Biol. 2023. PMID: 37400509 Free PMC article. Review.
Cited by
-
Ontogeny of aerial righting and wing flapping in juvenile birds.Biol Lett. 2014 Aug;10(8):20140497. doi: 10.1098/rsbl.2014.0497. Biol Lett. 2014. PMID: 25165451 Free PMC article.
-
The molecular evolution of feathers with direct evidence from fossils.Proc Natl Acad Sci U S A. 2019 Feb 19;116(8):3018-3023. doi: 10.1073/pnas.1815703116. Epub 2019 Jan 28. Proc Natl Acad Sci U S A. 2019. PMID: 30692253 Free PMC article.
-
Ontogenetic changes in the olfactory antennules of the shore crab, Hemigrapsus oregonensis, maintain sniffing function during growth.J R Soc Interface. 2015 Jan 6;12(102):20141077. doi: 10.1098/rsif.2014.1077. J R Soc Interface. 2015. PMID: 25411408 Free PMC article.
-
Intraskeletal histovariability, allometric growth patterns, and their functional implications in bird-like dinosaurs.Sci Rep. 2018 Jan 10;8(1):258. doi: 10.1038/s41598-017-18218-9. Sci Rep. 2018. PMID: 29321475 Free PMC article.
-
Mechanisms for Mid-Air Reorientation Using Tail Rotation in Gliding Geckos.Integr Comp Biol. 2021 Sep 8;61(2):478-490. doi: 10.1093/icb/icab132. Integr Comp Biol. 2021. PMID: 34143210 Free PMC article.
References
-
- Dudley R, Yanoviak SP (2011) Animal aloft: The origins of aerial behavior and flight. Integrative And Comparative Biology. - PubMed
-
- Xu X, Zhou Z, Wang X, Kuang X, Zhang F, et al. (2003) Four-winged dinosaurs from China. Nature 421: 335. - PubMed
-
- Zhou Z, Barret P, Hilton J (2003) An exceptionally preserved Lower Cretaceous ecosystem. Nature 421: 807–814. - PubMed
-
- Zhou Z, Zhang F (2006) Mesozoic birds of China: A synoptic review. Vertebrata PalAsiatica 44: 73–98.
-
- Xu X, Zhang F (2005) A new maniraptoran dinosaur from China with long feathers of the metatar- sus. Naturwissenschaften 92: 173–177. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
