

Isolation and biochemical characterization of a glucose dehydrogenase from a hay infusion metagenome
- PMID: 24454935
- PMCID: PMC3891874
- DOI: 10.1371/journal.pone.0085844
Isolation and biochemical characterization of a glucose dehydrogenase from a hay infusion metagenome
Abstract
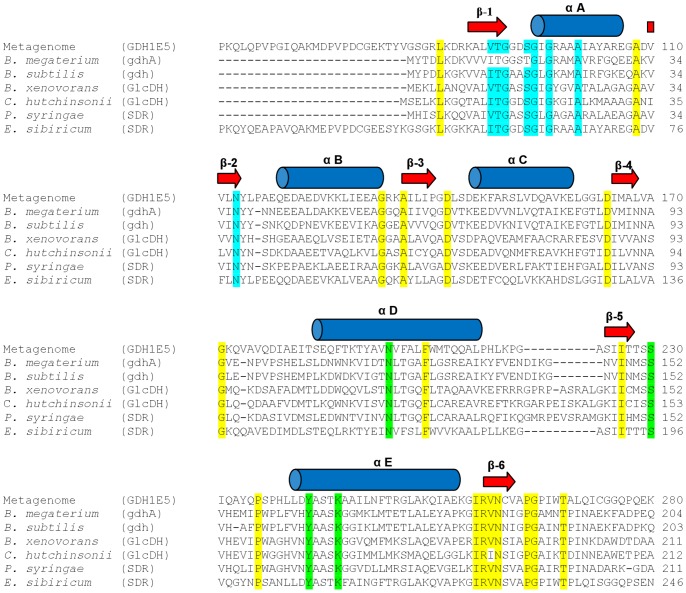
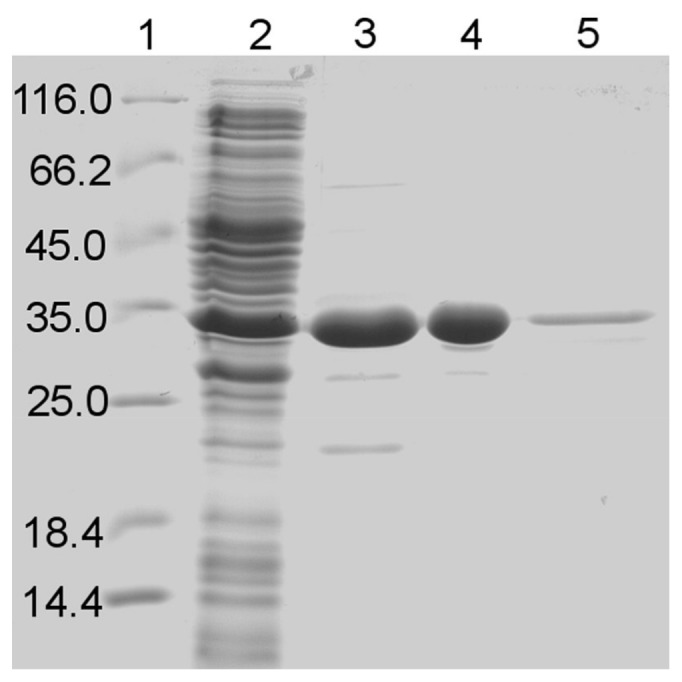
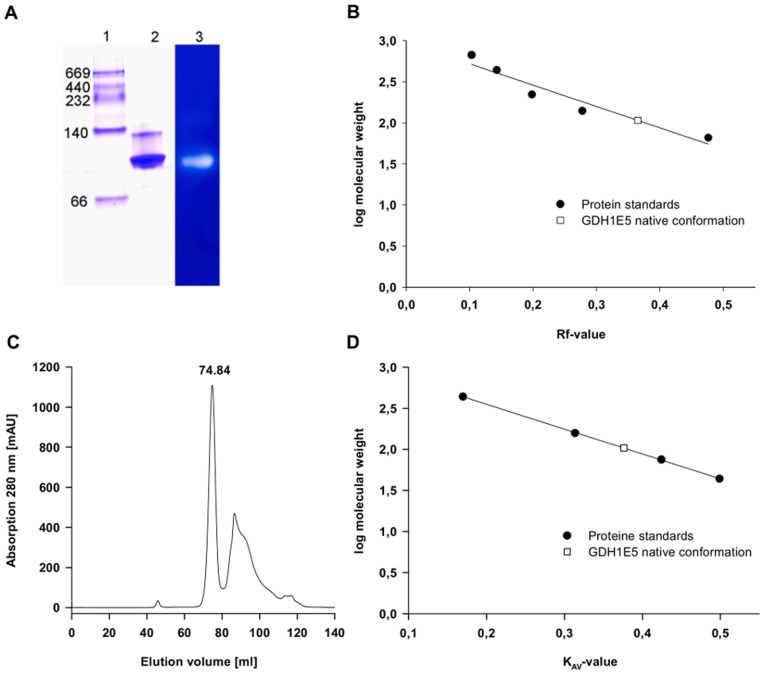
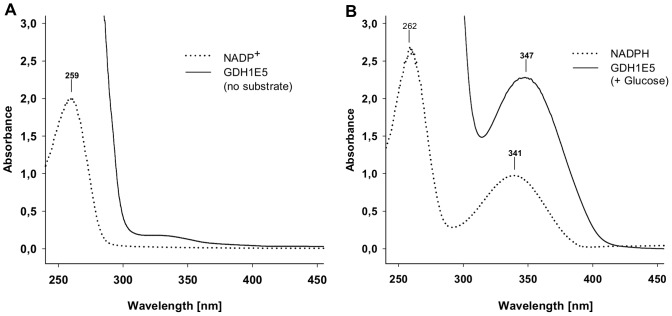
Glucose hydrolyzing enzymes are essential to determine blood glucose level. A high-throughput screening approach was established to identify NAD(P)-dependent glucose dehydrogenases for the application in test stripes and the respective blood glucose meters. In the current report a glucose hydrolyzing enzyme, derived from a metagenomic library by expressing recombinant DNA fragments isolated from hay infusion, was characterized. The recombinant clone showing activity on glucose as substrate exhibited an open reading frame of 987 bp encoding for a peptide of 328 amino acids. The isolated enzyme showed typical sequence motifs of short-chain-dehydrogenases using NAD(P) as a co-factor and had a sequence similarity between 33 and 35% to characterized glucose dehydrogenases from different Bacillus species. The identified glucose dehydrogenase gene was expressed in E. coli, purified and subsequently characterized. The enzyme, belonging to the superfamily of short-chain dehydrogenases, shows a broad substrate range with a high affinity to glucose, xylose and glucose-6-phosphate. Due to its ability to be strongly associated with its cofactor NAD(P), the enzyme is able to directly transfer electrons from glucose oxidation to external electron acceptors by regenerating the cofactor while being still associated to the protein.
Conflict of interest statement
Figures
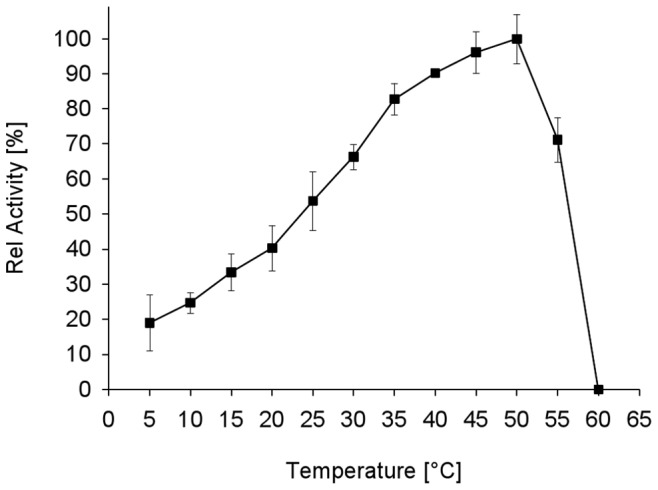
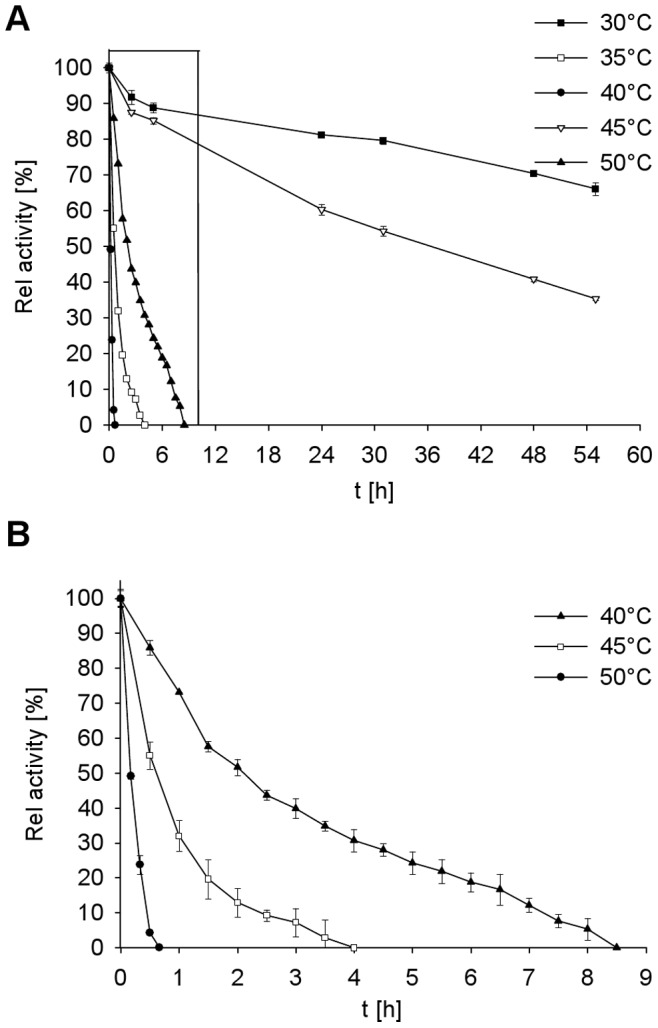
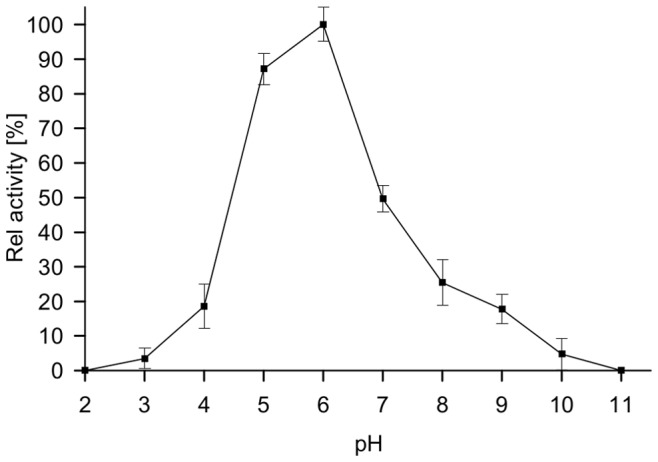
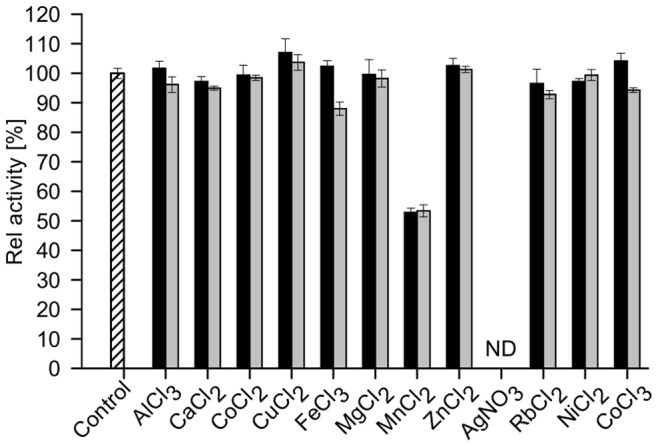
Similar articles
-
A novel glucose dehydrogenase from the white-rot fungus Pycnoporus cinnabarinus: production in Aspergillus niger and physicochemical characterization of the recombinant enzyme.Appl Microbiol Biotechnol. 2014 Dec;98(24):10105-18. doi: 10.1007/s00253-014-5891-4. Epub 2014 Jun 26. Appl Microbiol Biotechnol. 2014. PMID: 24965558
-
[Gene cloning and characterization of a solvent-resistant glucose dehydrogenase from Bacillus sp. YX-1].Wei Sheng Wu Xue Bao. 2013 Jun 4;53(6):561-8. Wei Sheng Wu Xue Bao. 2013. PMID: 24028058 Chinese.
-
Characterization of a thermostable glucose dehydrogenase with strict substrate specificity from a hyperthermophilic archaeon Thermoproteus sp. GDH-1.Biosci Biotechnol Biochem. 2015;79(7):1094-102. doi: 10.1080/09168451.2015.1018120. Epub 2015 Mar 9. Biosci Biotechnol Biochem. 2015. PMID: 25746627
-
Biochemical and Computational Insights on a Novel Acid-Resistant and Thermal-Stable Glucose 1-Dehydrogenase.Int J Mol Sci. 2017 Jun 5;18(6):1198. doi: 10.3390/ijms18061198. Int J Mol Sci. 2017. PMID: 28587256 Free PMC article.
-
Review of glucose oxidases and glucose dehydrogenases: a bird's eye view of glucose sensing enzymes.J Diabetes Sci Technol. 2011 Sep 1;5(5):1068-76. doi: 10.1177/193229681100500507. J Diabetes Sci Technol. 2011. PMID: 22027299 Free PMC article. Review.
Cited by
-
Functional analysis of BPSS2242 reveals its detoxification role in Burkholderia pseudomallei under salt stress.Sci Rep. 2020 Jun 26;10(1):10453. doi: 10.1038/s41598-020-67382-y. Sci Rep. 2020. PMID: 32591552 Free PMC article.
-
Prospects of advanced metagenomics and meta-omics in the investigation of phytomicrobiome to forecast beneficial and pathogenic response.Mol Biol Rep. 2022 Dec;49(12):12165-12179. doi: 10.1007/s11033-022-07936-7. Epub 2022 Sep 28. Mol Biol Rep. 2022. PMID: 36169892 Review.
-
One-step selective affinity purification and immobilization of His-tagged enzyme by recyclable magnetic nanoparticles.Eng Life Sci. 2021 May 4;21(6):364-373. doi: 10.1002/elsc.202000093. eCollection 2021 Jun. Eng Life Sci. 2021. PMID: 34140847 Free PMC article.
-
Obligately aerobic human gut microbe expresses an oxygen resistant tungsten-containing oxidoreductase for detoxifying gut aldehydes.Front Microbiol. 2022 Aug 16;13:965625. doi: 10.3389/fmicb.2022.965625. eCollection 2022. Front Microbiol. 2022. PMID: 36051760 Free PMC article.
-
Enzymes of an alternative pathway of glucose metabolism in obligate methanotrophs.Sci Rep. 2021 Apr 22;11(1):8795. doi: 10.1038/s41598-021-88202-x. Sci Rep. 2021. PMID: 33888823 Free PMC article.
References
-
- Anastassiadis S, Morgunov IG (2007) Gluconic acid production. Recent Pat Biotechnol 1 (2): 167–180. - PubMed
-
- Hönes J, Müller P, Surridge N (2008) The Technology Behind Glucose Meters: Test Stripes. DIABETES TECHNOLOGY & THERAPEUTICS (Volume 10): 10–26.
-
- Tsujimura S, Kojima S, Kano K, Ikeda T, Sato M, et al. (2006) Novel FAD-dependent glucose dehydrogenase for a dioxygen-insensitive glucose biosensor. Biosci. Biotechnol. Biochem. 70 (3): 654–659. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
