

Transcriptionally Repressive Chromatin Remodelling and CpG Methylation in the Presence of Expanded CTG-Repeats at the DM1 Locus
- PMID: 24455202
- PMCID: PMC3884603
- DOI: 10.1155/2013/567435
Transcriptionally Repressive Chromatin Remodelling and CpG Methylation in the Presence of Expanded CTG-Repeats at the DM1 Locus
Abstract
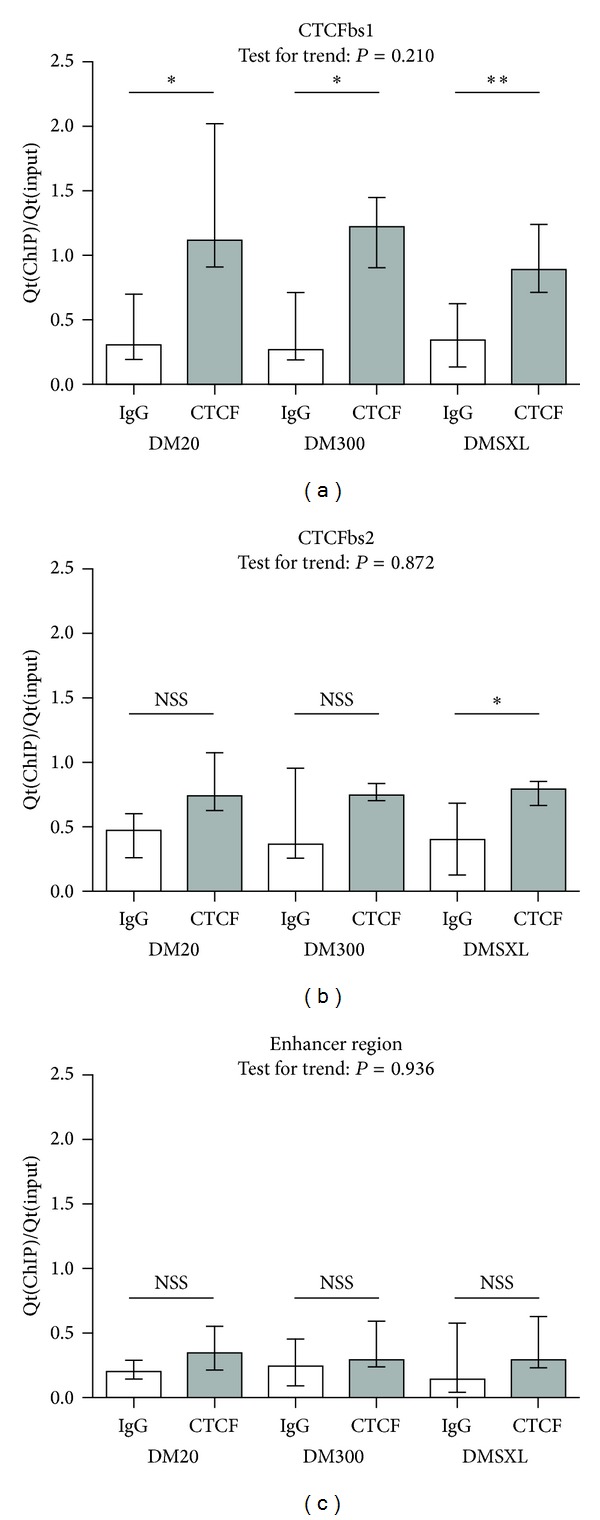
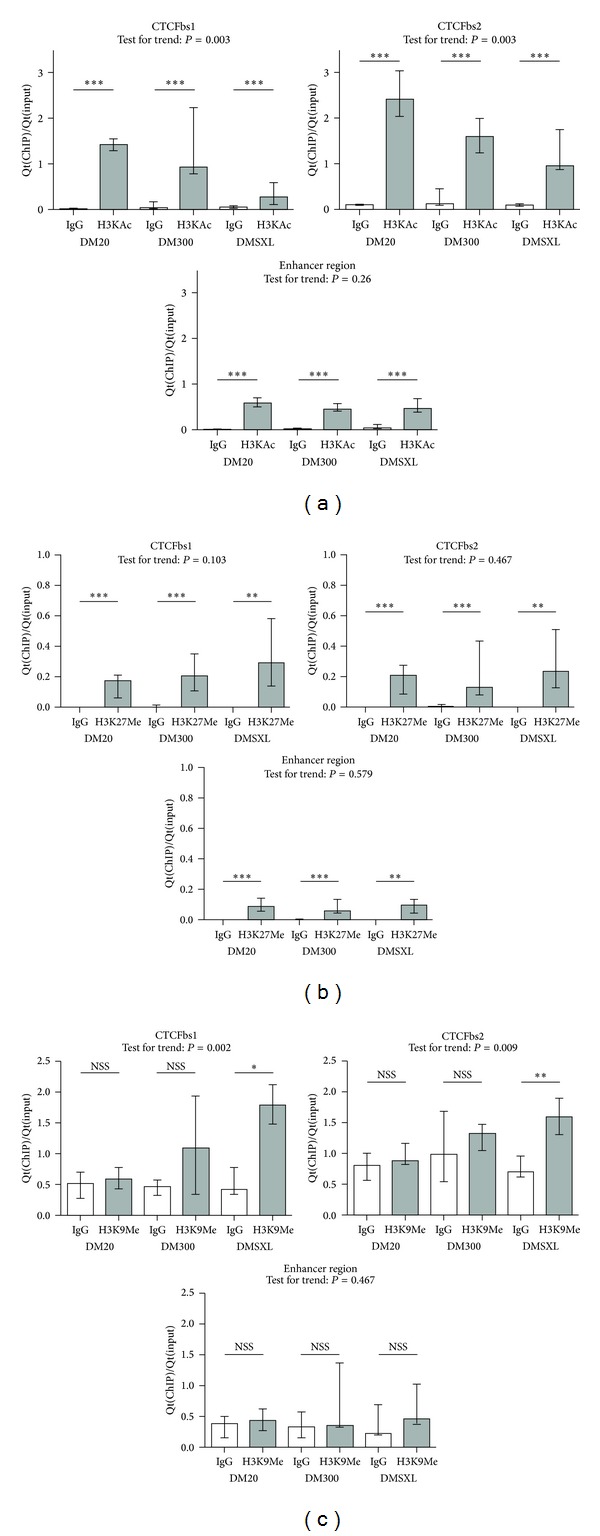
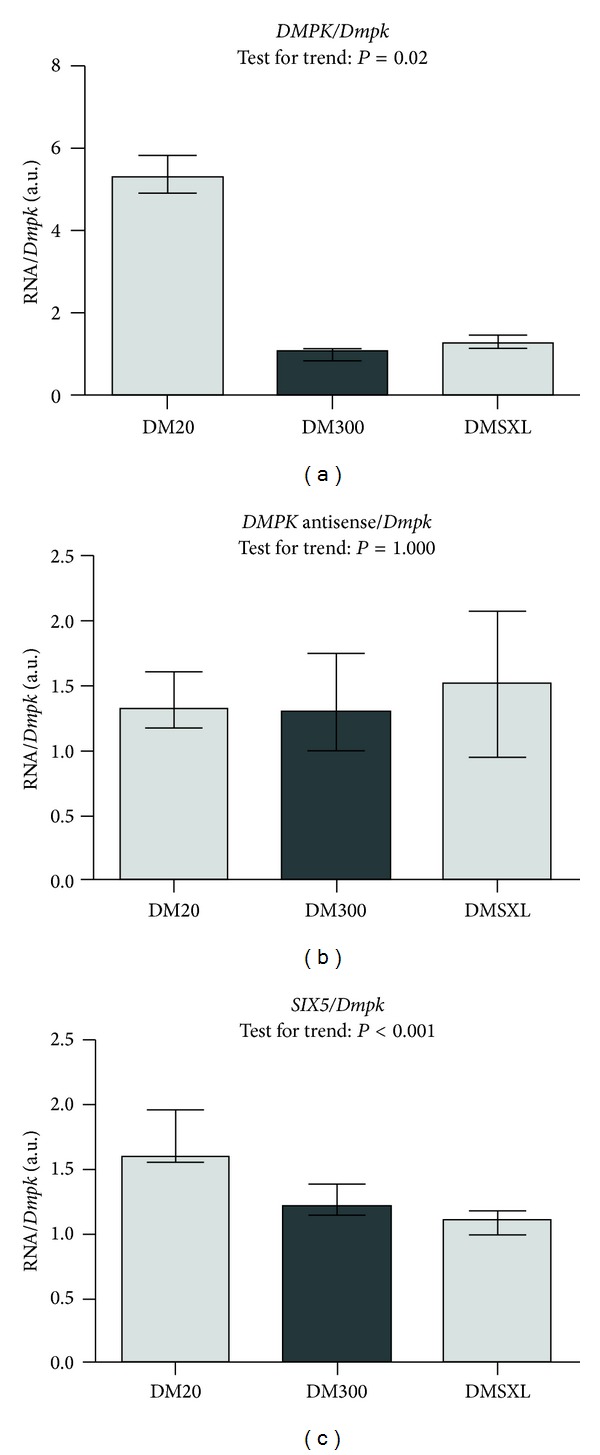
An expanded CTG-repeat in the 3' UTR of the DMPK gene is responsible for myotonic dystrophy type I (DM1). Somatic and intergenerational instability cause the disease to become more severe during life and in subsequent generations. Evidence is accumulating that trinucleotide repeat instability and disease progression involve aberrant chromatin dynamics. We explored the chromatin environment in relation to expanded CTG-repeat tracts in hearts from transgenic mice carrying the DM1 locus with different repeat lengths. Using bisulfite sequencing we detected abundant CpG methylation in the regions flanking the expanded CTG-repeat. CpG methylation was postulated to affect CTCF binding but we found that CTCF binding is not affected by CTG-repeat length in our transgenic mice. We detected significantly decreased DMPK sense and SIX5 transcript expression levels in mice with expanded CTG-repeats. Expression of the DM1 antisense transcript was barely affected by CTG-repeat expansion. In line with altered gene expression, ChIP studies revealed a locally less active chromatin conformation around the expanded CTG-repeat, namely, decreased enrichment of active histone mark H3K9/14Ac and increased H3K9Me3 enrichment (repressive chromatin mark). We also observed binding of PCNA around the repeats, a candidate that could launch chromatin remodelling cascades at expanded repeats, ultimately affecting gene transcription and repeat instability.
Figures



References
-
- Richards RI, Sutherland GR. Heritable unstable DNA sequences. Nature Genetics. 1992;1(1):7–9. - PubMed
-
- Aslanidis C, Jansen G, Amemiya C, et al. Cloning of the essential myotonic dystrophy region and mapping of the putative defect. Nature. 1992;355(6360):548–551. - PubMed
-
- Brook JD, McCurrach ME, Harley HG, et al. Molecular basis of myotonic dystrophy: expansion of a trinucleotide (CTG) repeat at the 3′ end of a transcript encoding a protein kinase family member. Cell. 1992;68(4):799–808. - PubMed
-
- Fu Y-H, Pizzuti A, Fenwick RG, Jr., et al. An unstable triplet repeat in a gene related to myotonic muscular dystrophy. Science. 1992;255(5049):1256–1258. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
